


ARN não codificador longo MALAT1 / microRNA-143 / Eixo de sinal VEGFA modula aneurisma intracraniano induzido por lesão endotelial vascular
Resumo
Os papéis de alguns longos RNAs não codificantes (lncRNAs) no aneurisma intracraniano (IA) foram investigados em muitos estudos. O objetivo deste estudo é elucidar o mecanismo do transcrito 1 de adenocarcinoma pulmonar associado à metástase de lncRNA (MALAT1) / microRNA-143 (miR-143) / fator de crescimento endotelial vascular-A (VEGFA) eixo de sinal em IA induzida por lesão endotelial vascular . A expressão de MALAT1, miR-143 e VEGFA em tecidos IA e tecidos arteriais normais foram detectados. Foram detectados metaloproteinase de matriz 9 (MMP-9) em tecidos, fator de von Willebrand (vWF) no soro e tecidos e endotelina-1 (ET-1) no soro. Os ratos IA modelados foram injetados com MALAT1 silenciado ou superexpresso para detectar lesão endotelial vascular. As células endoteliais vasculares de pacientes com IA foram abstraídas e transfectadas com MALAT1 silenciado ou superexpresso para verificar os impactos de MALAT1 na viabilidade celular e apoptose. As conexões entre MALAT1, miR-143 e VEGFA foram verificadas por predição online, atividade de luciferase e ensaios de pull down de RNA. A superexpressão de MALAT1 e VEGFA e a expressão pobre de miR-143 foram encontradas em tecidos IA. A regulação negativa de MALAT1 inibiu a pressão arterial, a expressão de ET-1, vWF e MMP-9, bem como o índice apoptótico de células endoteliais vasculares de ratos com IA. O MALAT1 regulado negativamente inibiu a apoptose e promoveu a viabilidade das células endoteliais vasculares em IA. MALAT1 ligado a miR-143 e miR-143 direcionados a VEGFA. Este estudo sugere que MALAT1 eleva a expressão de VEGFA por meio da ligação competitiva ao miR-143, aumentando assim a apoptose e atenuando a viabilidade das células endoteliais vasculares em IA.
Introdução
O aneurisma intracraniano (IA), também conhecido como aneurisma cerebral, é causado pelo aumento da pressão intracraniana induzida pelo alargamento anormal local da cavidade da artéria cerebral ou parede arterial [1]. IA é uma doença grave com alta mortalidade e morbidade, e a taxa de prevalência é de cerca de 1–3% na população em geral [2]. As principais características clínicas da IA são vasoespasmo cerebral, hemorragia cerebral espontânea e paralisia do nervo oculomotor [3]. Até agora, os fatores de risco comuns que levam à ocorrência e ao desenvolvimento de AI incluem distúrbios hemodinâmicos, genes, envelhecimento, infecções e fatores congênitos [4]. Os principais tratamentos clínicos, principalmente clipagem cirúrgica e / ou enrolamento endovascular, têm a função de prevenir a ruptura do aneurisma [5]. No entanto, o mecanismo detalhado subjacente à AI ainda precisa ser elucidado, refletindo a necessidade urgente de métodos mais eficazes no manejo da AI.
RNAs não codificantes longos (lncRNAs) têm mais de 200 nucleotídeos que pertencem a uma espécie de RNAs não codificantes [6]. É relatado que a expressão de lncRNA associado à parada de crescimento de lncRNA em IA é regulada negativamente e o transcrito de adenocarcinoma 1 (MALAT1) é um lncRNA altamente enriquecido e amplamente expresso cujo comprimento é de cerca de 8000 nt [7]. MALAT1 foi documentado para modular a disfunção do músculo liso no aneurisma da aorta torácica [8]. Além disso, um estudo apresentou a expressão anormal de lncRNAs e RNA mensageiro (mRNA) em IA, e as redes de coexpressão de lncRNA-mRNA fornecem pistas para encontrar a patogênese de IA [9]. Foi sugerido que MALAT1 avança a expressão de osterix e modula a diferenciação osteogênica por meio da expressão de miRNA-143 (miR-143) em células-tronco mesenquimais da medula óssea humana [10]. Um estudo também apresentou o papel do cluster miR-143/145 em reverter a regulação do fator 5 semelhante a krüppel nas células musculares lisas e sua contratilidade e proliferação em IA [11]. De acordo com Feng et al., A regulação negativa de miR-143/145 e nível superior de metaloproteinase-9 (MMP-9) durante a circulação pode estar relacionada à formação e ruptura de IA [12]. Uma análise revelou que a maioria dos miRNAs não controlados (miR-143 e miR-145) são comuns a genes-alvo que são vias de sinal, como fator de crescimento endotelial vascular (VEGF) e outros genes que regulam o movimento ou adesão celular [13] . Um estudo revelou a importância preditiva das variações do fator A de crescimento endotelial vascular (VEGFA) na IA [14]. Assim, buscamos avaliar o mecanismo do eixo de sinal MALAT1 / miR-143 / VEGFA em IA induzida por lesão endotelial vascular.
Materiais e métodos
Declaração de Ética
O estudo foi aprovado pelo Comitê de Revisão Institucional do Primeiro Hospital Afiliado da USTC, Divisão de Ciências da Vida e Medicina da Universidade de Ciência e Tecnologia da China e seguiu os princípios da Declaração de Helsinque. Os participantes forneceram consentimento informado por escrito neste estudo. Todos os experimentos com animais estavam de acordo com o Guia para o Cuidado e Uso de Animais de Laboratório do National Institutes of Health. O protocolo foi permitido pelo Comitê de Ética em Experimentos Animais do Primeiro Hospital Afiliado da USTC, Divisão de Ciências da Vida e Medicina da Universidade de Ciência e Tecnologia da China.
Assuntos de estudo
Vinte pacientes com IA (grupo IA) que foram diagnosticados por exame de imagem e receberam clipagem neurocirúrgica no The First Affiliated Hospital of USTC, Divisão de Ciências da Vida e Medicina da Universidade de Ciência e Tecnologia da China foram selecionados para nossos experimentos. Havia 11 homens e 9 mulheres com idades entre 43,27 ± 6,25 anos. Os tecidos IA foram amostrados. Enquanto isso, os tecidos vasculares corticais temporais do pólo temporal foram ressecados de 20 pacientes (grupo controle) com epilepsia do lobo temporal causada por amígdala e esclerose hipocampal. Os tecidos ressecados foram examinados como tecidos arteriais normais por histopatologia após a operação, e havia 13 homens e 7 mulheres com idades de 44,18 ± 5,91 anos. Nenhuma discrepância significativa foi reconhecida em gênero e idade entre o grupo IA e o grupo de controle (ambos P > 0,05). Amostras de sangue venoso (2 tubos) foram obtidas de todos os indivíduos em jejum ao mesmo tempo pela manhã antes da operação.
Estabelecimento de modelos de ratos de IA
Sessenta ratos machos Sprague-Dawley de grau limpo (Hunan SJA Laboratory Animal Co., Ltd., Hunan, China), pesando entre 200 e 250 g, foram criados por 7 dias (25 ± 2 ° C, umidade relativa de 65-70 %, 12 h de ciclo claro e escuro, ingestão de água e alimentos livres). Os ratos foram deixados cair com a elastase pancreática suína na artéria carótida externa e em torno da parede da artéria de bifurcação. A artéria carótida externa foi ligada com duas linhas cirúrgicas no ramo da artéria carótida externa cerca de 1,5 mm. A artéria carótida externa foi cortada entre as duas linhas para formar um aneurisma carotídeo interno no segmento cego da artéria carótida externa. Os ratos foram alimentados com solução salina a 1% durante 1 semana após a operação. A angiografia cerebral foi realizada após 1 mês e a formação de aneurisma foi observada.
Após o estabelecimento de modelos de ratos IA, 50 ratos foram distribuídos aleatoriamente em um grupo em branco ( n =10, os ratos modelados foram tratados com injeção estereotáxica de 100 μL de solução salina tamponada com fosfato (PBS) uma vez por dia), grupo de controle negativo (NC) de RNA em gancho curto (sh) ( n =10, os ratos modelados foram tratados com injeção estereotáxica de 100 μL sh-MALAT1 NC uma vez por dia), grupo sh-MALAT1 ( n =10, os ratos modelados foram tratados com injeção estereotáxica de 100 μL de plasmídeo sh-MALAT1 uma vez por dia), grupo de superexpressão (Oe) -NC ( n =10, os ratos modelados foram tratados com injeção estereotáxica de 100 μL de plasmídeo Oe-MALAT1 NC uma vez por dia) e grupo Oe-MALAT1 ( n =10, os ratos modelados foram tratados com injeção estereotáxica de 100 μL do plasmídeo Oe-MALAT1 uma vez por dia) [15]. Os plasmídeos acima foram compostos por Shanghai Genechem Co., Ltd. (Shanghai, China).
Teste de pressão arterial de ratos
A pressão arterial da artéria da cauda dos ratos foi medida na 1ª, 4ª e 12ª semanas após a operação. Antes de medir a pressão arterial, os ratos foram colocados em um dispositivo de aquecimento de temperatura constante por um momento para evitar a perturbação da temperatura externa. Em segundo lugar, os ratos foram mantidos em silêncio por vários minutos em uma gaiola especial para evitar a interferência da atividade. Se a pressão arterial variava muito, era determinada duas ou três vezes em momentos diferentes para obter o valor médio.
Aquisição de tecido de aneurisma
Após 3 meses, os ratos foram anestesiados com pentobarbital sódico a 1% (40 mg / kg) por injeção intraperitoneal para obter amostras de sangue das veias. Os ratos foram sacrificados e o tórax foi aberto, o ventrículo esquerdo foi entubado na aorta e a cava foi cortada para liberar o sangue. Enquanto isso, 30 mL de solução salina normal contendo heparina sódica foram utilizados para perfusão cardíaca rápida para limpar o sangue e, em seguida, 10 mL de poliformaldeído a 10% / PBS 0,1 M (pH 7,4) foram injetados no cérebro. Após perfusão e fixação, o cérebro do rato foi aberto. A circulação arterial na base do crânio foi cuidadosamente observada, o tecido do aneurisma foi separado e as alterações do aneurisma foram observadas ao microscópio.
Ensaio de imunoabsorção enzimática (ELISA)
Os índices relacionados ao soro foram testados pelo kit ELISA. As amostras de sangue coletadas foram colocadas em um termostato de 37 ° C por 1 he centrifugado a 3000 r / min por 10 min. A detecção da expressão da endotelina-1 (ET-1) e do fator de von Willebrand (vWF) foi conduzida de acordo com as instruções do kit (todos os kits foram adquiridos do NanJing JianCheng Bioengineering Institute, Jiangsu, China).
Coloração com Hematoxilina-Eosina (HE)
As amostras foram fixadas em formol a 10% por mais de 24 he conservadas em blocos de parafina. Os blocos de parafina foram desparafinados com xileno por 20 min, desidratados com gradiente descendente em série de álcool (100%, 95%, 80%, 75%) por l min e corados com hematoxilina por 10 min. Em seguida, os tecidos foram enxaguados com água destilada, diferenciados com ácido clorídrico etanol por 30 s, e embebidos em água morna a 50 ° C por 5 min. Tingidos com solução de eosina, os tecidos foram enxaguados com água destilada, desidratados com álcool 70% e 90%, clarificados com xilol e selados com goma neutra. A morfologia dos tecidos foi observada ao microscópio de alta potência.
Observação ao microscópio eletrônico de transmissão
Os tecidos sobressalentes foram fixados com dialdeído glutárico 2,5% e ácido osmítico 1%, desidratados e embebidos em resina Epon812. As seções semifinas foram tingidas com azul de tolueno, aparadas e confeccionadas em seções ultrafinas. As seções foram tingidas com acetato de uranila e citrato de chumbo e observadas por um microscópio eletrônico de transmissão JME-2000EX (Hitachi High-Technologies Co., Ltd., Shanghai, China).
Coloração terminal de desoxinucleotidil transferase mediada por desoxiuridina trifosfato-biotina Nick (TUNEL) coloração final
A coloração TUNEL foi sugerida para observar a apoptose das células endoteliais em ratos IA com base no kit TUNEL (Roche, Basel, Suíça). As seções de aneurisma de rato preparadas foram lavadas duas vezes com xileno (5 min / tempo) e desidratadas com séries descendentes de álcool (100%, 95%, 80%, 75%) por 3 vezes (5 min / tempo). Os tecidos foram tratados com 20 μg / mL de solução de protease K sem DNase por 15-30 min, descartados com 50 μL de solução de reação TUNEL por 60 min e desenvolvidos com 50 μL de diaminobenzidina (DAB) a 25 ° C por 10 min. Em seguida, os cortes foram contrastados com hematoxilina, desidratados com álcool gradiente, clarificados com xilol e selados com goma neutra. Os cortes foram observados ao microscópio óptico e o índice apoptótico calculado.
Isolamento e identificação de células endoteliais vasculares de aneurisma
O isolamento das células endoteliais foi realizado de acordo com o método conduzido por Boscolo et al. [16]. Os tecidos IA foram seccionados em 3 mm 2 fragmentos. Os tecidos foram incubados com 0,1% de colagenase B / 0,1% de dispase (Roche) durante 25 min a 37 ° C. Em seguida, os tecidos pré-destacados foram triturados por 2 min por uma pipeta de 2 mL e filtrados com uma peneira de 100 μm (Thermo Fisher Scientific, Rockford, IL, EUA). Posteriormente, a suspensão de células foi centrifugada e, em seguida, ressuspensa em meio MV2 (incluindo fatores de crescimento e 20% de soro fetal bovino) (PromoCell, Heidelberg, Alemanha). As células foram semeadas em 1 × 10 4 células / mL em um frasco de cultura revestido com 1 μg / cm 2 fibronectina. Seguindo o método descrito por Jackson et al. [17], as células de 80-100% de confluência foram separadas com as contas (Dynabeads M-450 Tosylactived, Oxoid, Hampshire, UK) revestidas por Ulex europaeus Agglutinin I (UEA) (Vector Laboratories, Ltd., Peterborough, UK) . As células endoteliais aderidas às esferas revestidas com lectina foram acumuladas com um concentrador de partículas magnéticas e as células não conjugadas foram lavadas com um meio basal. As células positivas para UEA foram ressuspensas no meio de cultura e semeadas no frasco de cultura revestido com fibronectina para melhorar a adesão e a taxa de crescimento das células.
As células foram cultivadas em MV2 em lâminas de câmara revestidas com fibronectina. Quando a confluência celular atingiu 80-100%, as células foram fixadas em acetona a 4 ° C e tratadas com 1% Triton X-100 por 5 min e, em seguida, 0,5% de albumina sérica bovina (BSA) por 15 min. As células foram gotejadas com anticorpo primário contra vWF (1:300, Abcam, Cambridge, MA, EUA) e incubadas por 2 h (NC foi realizado na ausência de anticorpo primário), gotejado com imunoglobulina G conjecturada com peroxidase de rábano (1:150, Abcam) e incubado durante 30 min. Em seguida, as células foram desenvolvidas com 50 μL de DAB a 25 ° C por 5 min, contracoradas com hematoxilina, diferenciadas com ácido clorídrico 0,1%, desidratadas com álcool, seguido de depuração de xileno e selamento de goma neutra. Após a secagem, as células foram fotografadas em microscópio invertido.
Agrupamento e transfecção de células
As células endoteliais vasculares de aneurisma na fase logarítmica foram distribuídas em 5 grupos:grupo em branco (células endoteliais vasculares de aneurisma sem qualquer tratamento), grupo sh-NC (células endoteliais vasculares de aneurisma transfectadas com plasmídeo sh-MALAT1 NC), grupo sh-MALAT1 ( células endoteliais vasculares de aneurisma transfectadas com plasmídeo sh-MALAT1), grupo Oe-NC (células endoteliais vasculares de aneurisma transfectadas com plasmídeo Oe-MALAT1 NC) e grupo Oe-MALAT1 (células endoteliais vasculares de aneurisma transfectadas com plasmídeo Oe-MALAT1). Os plasmídeos acima foram sintetizados por Genechem. A transfecção celular foi realizada de acordo com as instruções da lipofectamina TM Reagente 2000 (11668-027, Invitrogen, Carlsbad, Califórnia, EUA).
Ensaio de 3- (4, 5-Dimetiltiazol-2-Yl) -2, 5-difeniltetrazólio (MTT)
As células endoteliais vasculares de cada grupo foram semeadas em uma placa de 96 poços a uma densidade de 3 × 10 4 células / mL e cultivadas a 37 ° C, 5% CO 2 por 48 h. Cada grupo foi definido com 5 poços paralelos, e cada poço foi anexado com 20 μL de solução de MTT fresca (5 mg / mL, Sigma, St. Louis, MO, EUA). Após 4 h de reação, as células foram misturadas com 200 μL de dimetilsulfóxido. Após a dissolução total, o valor da densidade óptica das células em cada grupo foi medido por um leitor de microplacas (BioRad, Hercules, Califórnia, EUA) a 490 nm.
Citometria de fluxo
A distribuição do ciclo celular foi testada por coloração com iodeto de propídio (PI). As células endoteliais vasculares foram destacadas, centrifugadas, ressuspensas com etanol 75% pré-resfriado e fixadas durante a noite a -20 ° C. As células foram centrifugadas para descartar o sobrenadante. As células foram anexadas em 450 μL de PBS, adicionado com 100 μL de Rnase A e coradas por 400 μL de PI a 4 ° C por 30 min, evitando a luz. Um citômetro de fluxo (FACSCalibur, Becton, Dickinson and Company, NJ, EUA) foi utilizado para testar a distribuição do ciclo celular.
A apoptose celular foi testada por coloração dupla com Anexina V / PI. As células endoteliais destacadas foram reunidas e lavadas com PBS 3 vezes. As células foram ressuspensas com 100 μL de tampão de ligação 1 × pré-resfriado e misturadas com 5 μL de anexina e 5 μL de PI, respectivamente. A apoptose celular foi testada por um citômetro de fluxo. Com AnnexinV como eixo transversal e PI como eixo longitudinal, o quadrante superior esquerdo representava células danificadas mecanicamente (AnnexinV - / PI + ), o quadrante superior direito para células apoptóticas tardias ou células necróticas (AnexinV + / PI + ), o quadrante inferior esquerdo para células normais negativas (AnnexinV - / PI - ), e o quadrante inferior direito para células apoptóticas iniciais (AnnexinV + / PI - ) O total de células apoptóticas (AnnexinV + / PI - e AnnexinV + / PI + ) foram calculados e expressos como uma porcentagem.
Reação em cadeia da polimerase quantitativa por transcrição reversa (RT-qPCR)
Os RNAs totais nos tecidos e células foram abstraídos por Trizol (Takara Biotechnology Ltd., Dalian, China), e a concentração e pureza do RNA foram determinadas. O processo de transcrição reversa do RNA em DNA complementar foi realizado de acordo com as instruções do kit de transcrição reversa (K1621, Fermentas, Maryland, NY, USA). As sequências dos iniciadores MALAT1, miR-143 e VEGFA (Tabela 1) foram idealizadas e compostas pela Genechem. Para avaliar a expressão de lncRNA, miRNA ou mRNA, RT-qPCR foi conduzido usando SYBR GreenPCR Master Mix (Takara, Tóquio, Japão) com o sistema Roche Real-Time PCR (Roche). U6 foi definido como um parâmetro interno de miR-143, enquanto MALAT1 e VEGFA, com gliceraldeído-3-fosfato desidrogenase (GAPDH) como parâmetros internos. Os níveis transcricionais relativos dos genes alvo foram calculados por 2 - △△ Ct método.
Análise de Western Blot
A proteína total dos tecidos e células foi abstraída. A concentração de proteína foi determinada pelas instruções do kit de ácido bicinconínico (Boster Biological Technology Co. Ltd., Wuhan, Hubei, China). A proteína foi separada por eletroforese em gel de poliacrilamida 10% (Boster Biological Technology). A membrana foi transferida para uma membrana de fluoreto de polivinilideno e então selada com 5% BSA por 1 h. A membrana foi incubada com anticorpo primário contra Ki-67 (1:1000), VEGFA (1:1000), vWF (1:1000) e metaloproteinase de matriz (MMP) -9 (1:1000, Abcam, Cambridge, UK) , Cyclin D1 (1:1000), Bax (1:1000) e Bcl-2 (1:1000, Santa Cruz Biotechnology, Santa Cruz, Califórnia, EUA) e GAPDH (1:2000, Jackson Immuno Research, Grove, Pensilvânia, EUA) e com o anticorpo secundário (1:500, Jackson Immuno Research) marcado com peróxido de rábano. A membrana foi obtida pelo sistema de imagem de varredura por fluorescência de infravermelho de duas cores Odyssey, e os valores de cinza das bandas foram medidos pelo software de análise de imagem Quantity One.
Ensaio do gene Dual Luciferase Reporter
Os sítios de ligação de MALAT1 e miR-143 foram previstos e expostos pelo site de bioinformática (https://cm.jefferson.edu/rna22/Precomputed/). A relação de ligação entre MALAT1 e miR-143 foi ainda verificada pelo ensaio do gene repórter da luciferase. O fragmento do gene sintético da região 3′ não traduzida de MALAT1 (3′UTR) foi introduzido em vetores repórter de luciferase pmiR-Report (Thermo Fisher Scientific) para gerar MALAT1 de tipo selvagem (MALAT1-WT) pelo sítio de endonuclease Bamh1 e Ecor1. O local de mutação da sequência complementar da sequência foi desenvolvido em MALAT1-WT e o fragmento alvo foi inserido nos vetores repórter de luciferase pmiR-Report para produzir MALAT1 tipo mutante (MALAT1-MUT) por T4 DNA ligase após digestão com endonuclease de restrição. Os MALAT1-WT e MALAT1-MUT sequenciados corretamente foram co-transfectados com mímico NC e miR-143 mímico em células endoteliais vasculares. As células foram colhidas e lisadas 48 h após a transfecção, e a atividade da luciferase foi testada pelo kit de detecção de luciferase (BioVision, San Francisco, CA, EUA) com um luminômetro (Glomax20 / 20, Promega, Madison, Wisconsin, EUA).
A relação alvo de miR-143 e VEGFA e o sítio de ligação de miR-143 e VEGFA 3′UTR foram previstos por meio do site de bioinformática (http://www.targetscan.org/vert_72/). A sequência da região do promotor VEGFA 3′UTR contendo o sítio de ligação miR-143 foi composta e clonada em vetores repórter de luciferase pmiR-Report para gerar VEGFA-WT. Com base neste repórter, o sítio de ligação de VEGFA-WT e miR-143 foi mutado para formar VEGFA-MUT. O repórter VEGFA-WT ou VEGFA-MUT foi misturado com o imitador NC ou miR-143 imitador e, em seguida, co-transfectado em células endoteliais vasculares durante 48 h. Depois disso, as células foram lisadas e a atividade da luciferase foi testada por um kit de detecção de luciferase.
Ensaio RNA-pull Down
Para verificar a relação de ligação entre miR-143 e MALAT1, o RNA-pull down assay foi implementado. Três sequências de miRNA marcadas com biotina Bio-miR-143-WT, Bio-miR-143-MUT e Bio-miR-NC foram projetadas e confiadas à GenePharma Company (Shanghai, China). Estes oligonucleotídeos biotinilados foram transfectados em células por 48 h. Em seguida, as células foram colhidas e incubadas com um lisado celular específico (Ambion, Austin, Texas, EUA) por 10 min. Depois disso, o lisado foi incubado com esferas de estreptavidina M-280 pré-revestidas com tRNA livre de RNase e de levedura (todos da Sigma) a 4 ° C por 3 h, em seguida, limpo duas vezes com uma solução de lise fria, três vezes com um baixo tampão salino e uma vez com solução tampão com alto teor de sal. Uma sonda miR-143 antagonista foi estabelecida como NC. O RNA total foi abstraído com Trizol e, em seguida, o nível de enriquecimento MALAT1 foi testado usando RT-qPCR.
Análise estatística
Todos os dados foram expostos pelo software SPSS 21.0 (IBM Corp. Armonk, NY, EUA). Os dados de medição foram representados por média ± desvio padrão. As comparações entre dois grupos foram realizadas por amostra independente t teste, enquanto as comparações entre vários grupos foram avaliadas por análise de variância unilateral (ANOVA), e a comparação de pares foi implementada pelo teste de comparações múltiplas de Tukey. A relação entre a expressão de MALAT1 e as características clínico-patológicas de IA foi determinada pelo teste do qui-quadrado. P valor inferior a 0,05 foi indicativo de diferença estatisticamente significativa.
Resultados
MALAT1 e VEGFA são superexpressos e miR-143 é regulado para baixo em tecidos IA
A expressão de ET-1 e vWF no soro no grupo IA e no grupo de controle foi detectada por ELISA, e os resultados manifestaram que a expressão de ET-1 e vWF aumentou no grupo IA em relação ao grupo controle (ambos P <0,05) (Fig. 1a, b).
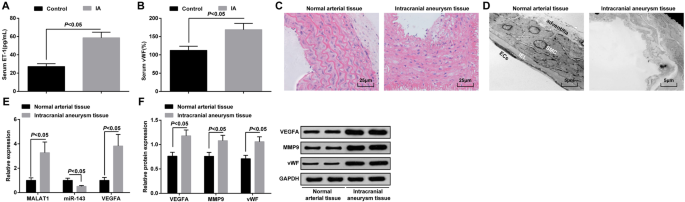
MALAT1 e VEGFA são superexpressos e miR-143 é regulado negativamente em tecidos IA. a Expressão de ET-1 no soro de pacientes com IA e pacientes com epilepsia do lobo temporal por ELISA. b Expressão do vWF no soro de pacientes com IA e pacientes com epilepsia do lobo temporal por ELISA. c Observação patológica de tecidos IA e tecidos arteriais normais por coloração HE. d Observação morfológica de tecidos IA e tecidos arteriais normais por um microscópio eletrônico de transmissão. e Expressão de mRNA de MALAT1, miR-143 e VEGFA em tecidos IA e tecidos arteriais normais por RT-qPCR. f Expressão de proteínas de VEGFA, MMP-9 e vWF em tecidos IA e tecidos arteriais normais por análise de Western blot. Células endoteliais (CEs), lâmina elástica interna (IEL), células musculares lisas (SMC). Os dados de medição foram representados como média ± desvio padrão; as comparações entre os grupos foram realizadas por amostra independente t teste
As alterações patológicas dos tecidos IA foram observadas através da coloração HE. Nenhum dano óbvio no endoderma, células endoteliais e células musculares lisas foi visto em tecidos arteriais normais, e as células foram organizadas ordenadamente e tinham uma estrutura completa. Os tecidos IA apresentavam células endoteliais danificadas, células musculares lisas degeneradas, parede arterial atenuada, fibras elásticas rompidas e células inflamatórias infiltradas (fig. 1c).
Alterações na ultraestrutura de IA e tecidos arteriais normais foram observadas por um microscópio eletrônico de transmissão. Verificou-se que as células endoteliais estavam completas e a estrutura da adventícia intacta; nenhuma membrana elástica interna quebrada ou células de músculo liso apoptóticas foram vistas em tecidos arteriais normais. Em tecidos IA, apresentou degeneração severa da parede dos vasos sanguíneos, manifestada principalmente como o desaparecimento da maioria das células endoteliais, camada elástica interna severamente rompida, células musculares lisas severamente danificadas e degradadas e o distúrbio da membrana externa do vaso sanguíneo (Fig. 1d).
RT-qPCR foi conduzido para determinar a expressão de MALAT1, miR-143 e VEGFA mRNA, enquanto a análise de western blot para VEGFA, MMP-9 e expressão da proteína vWF em tecidos IA e tecidos arteriais normais. Foi demonstrado que, em contraste com os tecidos arteriais normais, os níveis de expressão de MALAT1, VEGFA, MMP-9 e vWF foram aumentados e a expressão de miR-143 foi degradada em tecidos IA (todos P <0,05) (Fig. 1e, f).
Grau de Hunt-Hess, grau de dano endotelial e histórico de tabagismo estão correlacionados com a expressão de MALAT1 em tecidos IA
À luz da expressão mediana de MALAT1, os pacientes foram divididos em dois grupos:grupo de baixa expressão e grupo de alta expressão. A relação entre a expressão de MALAT1 e as características clínico-patológicas de pacientes com IA foi analisada. Os resultados sugeriram que o grau de Hunt-Hess, o grau de dano endotelial e a história de tabagismo foram associados à expressão de MALAT1 (todos P <0,05), enquanto idade, sexo e modo cirúrgico não foram relacionados à expressão de MALAT1 (todos P > 0,05) (Tabela 2).
MALAT1 regulado negativamente reprime a pressão arterial, a expressão de ET-1, vWF e MMP-9, bem como o índice apoptótico de células endoteliais vasculares de ratos com IA
Conforme exibido na Tabela 3, o knockdown de MALAT1 degradou-se, enquanto a restauração de MALAT1 aumentou a pressão arterial na 4ª e 12ª semanas.
O ELISA revelou que a regulação negativa de MALAT1 reduziu, enquanto a regulação positiva de MALAT1 elevou a expressão de ET-1 e vWF no soro de ratos com IA (Fig. 2a, b).
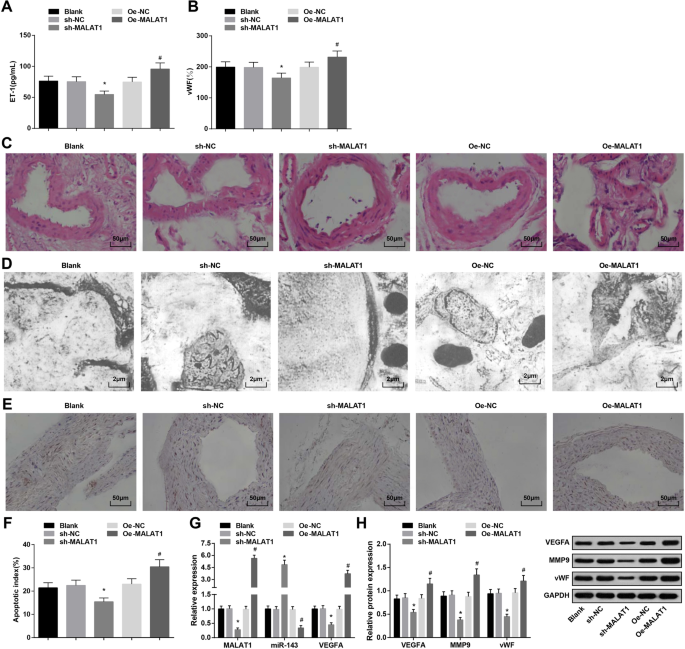
O MALAT1 regulado para baixo reprime a pressão arterial, a expressão de ET-1, vWF e MMP-9, bem como o índice apoptótico de células endoteliais vasculares de ratos com IA. a Expressão de ET-1 em soro de ratos por ELISA. b Expressão do vWF em soro de ratos por ELISA. c Alterações patológicas de tecidos IA em ratos observadas por coloração HE. d A ultraestrutura dos tecidos IA em ratos observada por um microscópio eletrônico de transmissão. e Apoptose de células endoteliais vasculares por coloração TUNEL. f Índice apoptótico de células endoteliais vasculares de ratos. g Expressão de mRNA de MALAT1, miR-143 e VEGFA em tecidos IA de ratos por RT-qPCR. h Expressão de proteínas de VEGFA, MMP-9 e vWF em tecidos IA de ratos por análise de Western blot. * P <0,05 vs. o grupo sh-NC, # P <0,05 vs. o grupo Oe-NC. Os dados de medição foram descritos como média ± desvio padrão, e as comparações entre vários grupos foram avaliadas por uma análise de variância unilateral seguida do teste de comparações múltiplas de Tukey
As alterações patológicas dos tecidos IA em cada grupo foram pesquisadas com coloração HE. O grupo em branco, o grupo sh-NC e o grupo Oe-NC foram manifestados com membrana interna danificada, células endoteliais esfoliadas, células musculares lisas degeneradas, células e camadas reduzidas e parede arterial mais fina. No grupo sh-MALAT1, a endoderme, as células endoteliais, a camada de células do músculo liso e a camada de membrana externa da artéria intracraniana foram levemente danificadas, mas organizadas de maneira organizada. O grupo Oe-MALAT1 apresentou desaparecimento da camada íntima, células endoteliais esfoliadas, fibras elásticas rompidas e células inflamatórias infiltradas (fig. 2c).
Os tecidos IA de ratos em cada grupo foram observados por um microscópio eletrônico de transmissão. Foi demonstrado que as células endoteliais desnaturadas, a camada subendotelial desintegrada, a camada elástica interna desapareceu e as células musculares lisas diminuídas foram apresentadas no grupo em branco, grupo sh-NC e grupo Oe-NC. O grupo sh-MALAT1 demonstrou células endoteliais planas, núcleo oval e aumento das fibras de colágeno, mas sem camada elástica. O grupo Oe-MALAT1 foi caracterizado pelo desaparecimento das células endoteliais e separação da camada elástica da camada muscular, que foi desintegrada no lúmen (fig. 2d).
O índice apoptótico de células endoteliais vasculares em ratos IA foi testado por coloração TUNEL. O silenciamento de MALAT1 reduziu o índice apoptótico de células endoteliais vasculares, enquanto a superexpressão de MALAT1 exibiu o efeito oposto (Fig. 2e, f).
A detecção RT-qPCR de MALAT1, miR-143 e expressão de mRNA de VEGFA e a análise de western blot da expressão de proteínas de VEGFA, MMP-9 e vWF em tecidos IA apresentaram que a eliminação de MALAT1 deprimiu a expressão de MALAT1, VEGFA, MMP-9 e vWF e expressão miR-143 aumentada. Ao contrário, a elevação de MALAT1 impôs influências opostas sobre essas expressões gênicas (Fig. 2g, h).
Baixa expressão de MALAT1 aumenta a viabilidade e restringe a apoptose de células endoteliais vasculares em IA
A coloração imunohistoquímica foi empregada para detectar a expressão do marcador específico do endotélio vWF. Foi manifestado que o citoplasma das células endoteliais vasculares IA estava recoberto por finas partículas castanhas, o que era positivo, enquanto o citoplasma do seu grupo NC não apresentava partículas castanhas. Os resultados confirmaram que as células cultivadas eram células endoteliais (Fig. 3a, b).
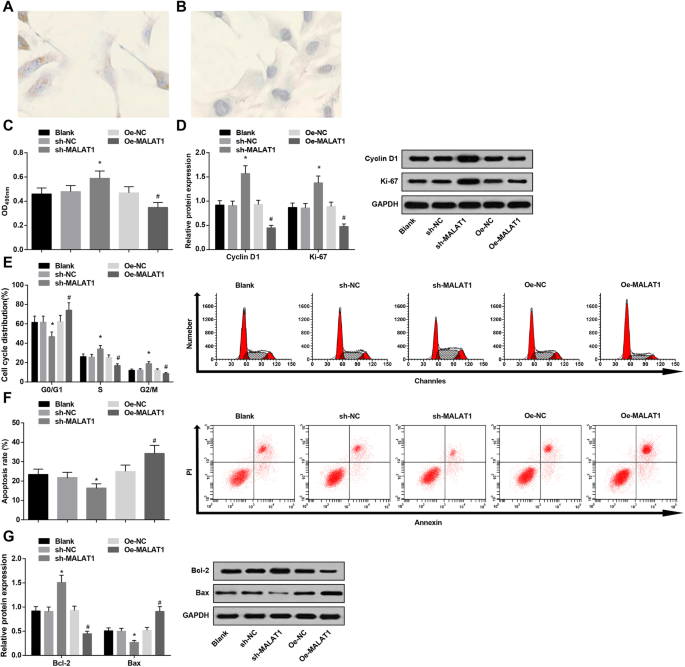
A baixa expressão de MALAT1 aumenta a viabilidade e restringe a apoptose de células endoteliais vasculares em IA. a vWF immunohistochemical staining in IA vascular endothelial cells:IA vascular endothelial cells were covered with fine yellow particles. b vWF immunohistochemical staining in IA vascular endothelial cells:IA vascular endothelial cells showed no brown particles in the NC group. c Vascular endothelial cell viability in each group by MTT assay. d Protein expression of CyclinD1 and Ki-67 in each group by western blot analysis. e Cell cycle changes in each group by PI staining. f Cell apoptosis rate in each group by Annexin V/PI double staining. g Bax and Bcl-2 protein expression in each group by western blot analysis. * P <0.05 vs. the sh-NC group, # P <0.05 vs. the Oe-NC group. Measurement data were depicted as mean ± standard deviation, and comparisons among multiple groups were assessed by one-way analysis of variance followed with Tukey’s multiple comparisons test
MTT assay, flow cytometry, together with western blot analysis were utilized to test vascular endothelial cell viability and apoptosis. It was displayed that MALAT1 diminution promoted vascular endothelial cell viability (heightened Cyclin D1 and Ki-67 expression) and depressed apoptosis (decreased G0/G1 phase cells and increased S and G2/M phase cells, reduced Bax and elevated Bcl-2 expression). However, MALAT1 upregulation functioned in an opposite way to that of MALAT1 diminution on cell viability and apoptosis (Fig. 3c–g).
MiR-143 Is Bound to MALAT1 and VEGFA Is a Target Gene of miR-143
MALA1, miR-143, and VEGFA expression in vascular endothelial cells of each group were verified through RT-qPCR and western blot analysis. The results presented that MALA1 knockdown depressed MALA1 and VEGFA expression and enhanced miR-143 expression. On the contrary, MALA1 upregulation led to the increment in MALAT1 and VEGFA expression and the reduction in miR-143 expression (Fig. 4a, b).

MiR-143 is bound to MALAT1 and VEGFA is a target gene of miR-143. a MALAT1, miR-143, and VEGFA mRNA expression in vascular endothelial cells of aneurysm in each group. b VEGFA protein expression in vascular endothelial cells of aneurysm in each group. c The binding sites of MALAT1 and miR-143 predicted by the bioinformatics website. d The regulatory relation of MALLA1 and miR-143 validated by dual luciferase reporter gene assay. e The binding relationship between MALAT1 and miR-143 verified by RNA-pull down assay. f The binding sites of miR-143 and VEGFA predicted by the bioinformatics website. g The regulatory relation of miR-143 and VEGFA validated by dual luciferase reporter gene assay. * P <0.05 vs. the sh-NC group, # P <0.05 vs. the Oe-NC group. Measurement data were depicted as mean ± standard deviation, comparisons between two groups were assessed by independent sample t test, and comparisons among multiple groups were assessed by one-way analysis of variance followed with Tukey’s multiple comparisons test
The specific binding region between MALAT1 and miR-143 was divined by online analysis software (Fig. 4c). The results of dual luciferase reporter gene assay revealed that the luciferase activity was impaired in the MALAT1-WT + miR-143 mimic group versus the MALAT1-WT + mimic-NC group (P <0,05). However, there was no distinct difference in luciferase activity in the MALAT1-MUT + miR-143 mimic group relative to that in the MALAT1-MUT + mimic-NC group (P> 0.05), indicating that miR-143 was specifically bound to MALAT1 (Fig. 4d). The results of RNA-pull down assay reported that the enrichment level of MALAT1 in the Bio-miR-143-WT group was heightened compared to the Bio-miR-NC group (P <0.05), but there was no distinct difference in MALAT1 enrichment level in the Bio-miR-143-MUT group (P> 0.05) (Fig. 4e).
Bioinformatics software divined a targeted relationship between miR-143 and VEGFA (Fig. 4f). The results of luciferase activity showed that the relative luciferase activity repressed after VEGFA-WT and miR-143 mimic co-transfected into vascular endothelial cells (P <0,05). However, the relative luciferase activity of vascular endothelial cells was not affected by co-transfection of VEGFA-MUT and miR-143-mimic (P> 0.05) (Fig. 4g). It was indicated that VEGFA was a direct target gene of miR-143.
Discussion
IA is a serious intracranial disease, which mainly leads to subarachnoid hemorrhage [18]. A previous study has demonstrated the involvements of lncRNA-related competitive endogenous RNA networks in IA [19]. Also, a recent study has provided a proof that functional polymorphism in miR-143/145 gene promoter region is connected with the risk of IA [20]. In a study conducted by Xu et al. , it is shown that overexpression of angiogenic factors, such as VEGFA, may be related to IA formation and rupture [21]. In order to explain the molecular mechanism of MALAT1 in IA, a series of assays have been conducted and the results revealed that IA induced by vascular endothelial injury was regulated by MALAT1/miR-143/VEGFA signal axis.
Firstly, our study has provided substantial evidence that MALAT1 and VEGFA are upregulated and miR-143 is downregulated in IA tissues. A recent study has presented that MALAT1 is one of the most upregulated lncRNAs in the process of cerebral ischemia [22]. Another study has presented that MALAT1 is upregulated in ovarian cancer cells and intends to participate in the processes of ovarian cancer cell apoptosis, migration, and proliferation [23]. A study concerning to the expression profile of unruptured and ruptured IA has demonstrated that the expression of angiogenic factors such as VEGFA is upregulated in ruptured aneurysm [21]. Moreover, a clinical study has presented that the miR-143/145 cluster of IA patients is downregulated compared to healthy subjects [11]. In addition, it is previously discussed that miR-143/145 takes part in various biological processes associated with aneurysm formation and is downregulated in patients with IA [20]. All these findings are more or less echoed with the previous exploration outcomes.
Except for the abovementioned findings, this study has also explored the functional role of MALAT1 in IA through gain-off-function and loss-of-function assays. It could be summarized that downregulation of MALAT1 reduced blood pressure, serum levels of ET-1, and vWF and MMP-9 expression in IA tissues. It has been suggested previously that downregulation of MALAT1 restrains the upregulation of the glucose-induced ET-1 transcription product [24]. Also, it is reported that ectopic MALAT1 expression is the inducer of vWF generation [25]. Another study has verified that the depletion of MALAT1 in bone marrow-derived macrophages inhibits the expression of MMP-9 [26].
Also, cell experiments have been conducted for further confirmation of the functions of MALAT1 in IA. The results have suggested that MALAT1 knockdown promoted vascular endothelial cell viability and depressed apoptosis in IA. Similarly, it has been documented that disturbance of MALAT1 improves aortic mural architecture and retards aneurysm growth [8]. Supplementary to our study finding, there is research highlighting that poor expression of MALAT1 induces apoptosis and restrains proliferation of acute myeloid leukemia cells [27]. Another study has also demonstrated the inhibitory effects of MALAT1 knockdown on proliferation of human osteoarthritis cartilage cells [28]. Besides that, a prior research generally confirms that downregulation of MALAT1 can induce apoptosis and attenuate the proliferation of glioma cells [29]. Moreover, low expression of MALAT1 induced by RNA interference promotes apoptosis and suppresses proliferation of multiple myeloma cells [30]. Collectively, these studies have explained the molecular mechanism of MALAT1 in IA to some extent.
In addition, this study has evidenced that miR-143 is bound to MALAT1 and VEGFA is a target gene of miR-143. Similarly, a paper contends that MALAT1 binds directly to miR-143 and suppresses its expression [10]. Zhu et al. have found that MALAT1 exerts its roles via interacting with miR-143 in cervical carcinoma cells [31]. It is further confirmed that MALAT1 indirectly modulate VEGFA via miR-200b-3p [32]. Moreover, another study has suggested that miR-143-3p mediates ZFPM2 effect on a number of protein targets in blood, including VEGFA [33]. Nevertheless, the interactions among MALAT1, miR-143, and VEGFA in IA have not been discussed and need further study.
Conclusion
From these results, it is clear that MALAT1 knockdown depresses apoptosis and promotes viability of vascular endothelial cells in IA by modulating miR-143/VEGFA axis. The co-expression network suggests the connection between MALAT1 and miR-143 with the involvement of VEGFA. The findings in this study partially disclose the pathogenesis of IA initiation and progression, and the studied targets may be a notably potential entry point to reveal pathology of IA from another perspective. Limitedly, further large-scale studies are required to comprehensively illustrate the mechanisms of MALAT1/miR-143/VEGFA axis in IA.
Síntese de Nanopartículas de Ouro de Vários Tamanhos pelo Método de Redução Química com Diferente Polaridade de Solvente
Detector fotovoltaico infravermelho baseado na heterojunção p-GeTe / n-Si
Nanomateriais