


Potenciais efeitos osteoindutivos de nanopartículas de hidroxiapatita em células-tronco mesenquimais por interação celular endotelial
Resumo
A nano-hidroxiapatita (nano-HA) atraiu atenção substancial no campo da medicina regenerativa. As interações entre células endoteliais (CE) e células-tronco mesenquimais (MSC) são necessárias para a reconstrução óssea, mas a maneira pela qual o nano-HA interage neste processo permanece desconhecida. Aqui, investigamos a citotoxicidade e os efeitos osteoindutivos de nanopartículas de HA (HANPs) em MSCs usando um modelo de co-cultura indireta mediado por CEs e destacamos os mecanismos subjacentes. Verificou-se que em uma dose subcitotóxica, os HANPs aumentaram a viabilidade e expressão de genes de osteoblastos, bem como nódulos mineralizados e produção de fosfatase alcalina de CTMs. Esses fenômenos dependiam do HIF-1α secretado por CEs, que disparou a cascata de sinalização ERK1 / 2. Além disso, um modelo matemático de linhagem celular de dois estágios foi estabelecido para analisar quantitativamente o impacto do HIF-1α na diferenciação osteogênica de MSCs. Demonstrou que o HIF-1α exerceu um efeito estimulador dependente da dose na taxa de diferenciação osteogênica de MSCs até 1500 pg / mL, o que está de acordo com os resultados acima. Nossos dados indicam que as interações cooperativas entre HANPs, ECs e MSCs provavelmente servem para estimular a regeneração óssea. Além disso, o modelo de linhagem celular de dois estágios é um sistema in vitro útil para avaliar a influência potencial das moléculas efetoras na engenharia do tecido ósseo.
Introdução
A reconstrução de defeitos ósseos causados por trauma, malformação congênita ou ressecção cirúrgica representa um grande desafio para a cirurgia ortopédica [1]. A hidroxiapatita (HA), uma cerâmica bioativa representativa, foi empregada como substituto ósseo [2]. No entanto, propriedades mecânicas e osteoindutivas indesejáveis restringem sua aplicação clínica [3]. Nos últimos anos, o nano-HA demonstrou bioatividade melhor e mais otimizada e melhor desempenho mecânico devido às suas características biônicas únicas e atraiu um interesse significativo nos campos biomédicos relacionados à medicina regenerativa [4]. Quando o nano-HA é implantado em defeitos ósseos, várias células envolvidas no reparo ósseo serão expostas a ele. Como tal, é necessário avaliar o comportamento biológico do nano-HA. Várias linhas de evidência demonstraram diretamente que as nanopartículas de HA (HANPs) podem ser captadas por células-tronco mesenquimais derivadas de gelatina de Wharton humano (hWJ-MSCs) e células de osteoblastos do cordão umbilical humano, resultando em diferenciação osteogênica aprimorada [5,6,7]. Dua et al. relataram anteriormente a capacidade dos HANPs de promover a integração da cartilagem projetada na cartilagem de novo [8]; por outro lado, os HANPs inibem a capacidade angiogênica das células endoteliais da veia umbilical humana (HUVECs) [9]. Em termos de saúde humana, uma compreensão mais abrangente do impacto dos HANPs na regeneração óssea é garantida, e as aplicações contínuas de osso artificial baseado em nanopartículas aumentam a urgência de tais estudos.
A regeneração óssea é inevitavelmente acompanhada pela invasão de neovasos. Os CEs são o revestimento celular interno do sistema vascular que serve para fornecer sangue passivamente e também desempenha um papel na indução, especificação e orientação da regeneração de órgãos, bem como na manutenção da homeostase e do metabolismo [10, 11]. MSCs são uma parte do nicho periendotelial e possuem habilidades de auto-renovação e multi-diferenciação sob a indução de microambientes fisiológicos e bioquímicos particulares dentro de seus nichos residentes [12]. Tsai et al. descobriram que os CEs podem secretar endotelina-1 para direcionar as CTMs em direção à diferenciação das linhagens osteo e condro [13]. Além disso, Saleh et al. usaram análise de dados de microarray para identificar proteínas secretadas por HUVEC e vias de sinalização de crosstalk relacionadas que interagem com os receptores ligados à membrana MSC para aumentar a proliferação e diferenciação osteogênica [14]. Na engenharia de tecido ósseo, os HANPs podem entrar em contato com os neovasos e ser endocitados por CEs, que mostraram alterar a função fisiológica dessas células [9, 15]. Isso também pode influenciar as células osteoprogenitoras circundantes e afetar o reparo ósseo, alterando a sinalização parácrina. No entanto, embora o impacto direto dos HANPs nas MSCs tenha sido explorado, ainda falta um entendimento claro se os HANPs podem induzir indiretamente a diferenciação osteogênica de MSCs através de ECs, o que é essencial para nossa compreensão do efeito dos HANPs em relação para o reparo ósseo.
Neste estudo, em um esforço para obter mais informações sobre os efeitos biológicos de HANPs na interação entre CEs e MSCs, um modelo de co-cultura indireta foi estabelecido usando HUVECs e hWJ-MSCs. Ao utilizar este sistema, a citotoxicidade e os efeitos osteoindutivos de HANPs em hWJ-MSCs via sinalização parácrina mediada por HUVEC foram avaliados. Para identificar os principais fatores que influenciam as interações células endoteliais induzidas por HANP-MSC, os fatores solúveis no sobrenadante de HUVECs que foram estimulados com HANPs foram avaliados com ênfase nos mecanismos relacionados nos níveis de gene e proteína. Os resultados demonstraram que o fator indutível por hipóxia (HIF) -1α desempenha um papel crítico nessas interações.
Para observar quantitativamente e prever o impacto do HIF-1α no processo de osteogênese, foi estabelecido um modelo matemático que combina uma linhagem celular de dois estágios com o HIF-1α. Aqui, ao analisar os dados empíricos, o modelo de linhagem celular de dois estágios foi usado para prever o número de MSC e o grau de diferenciação em qualquer ponto de tempo, com base na densidade de semeadura celular inicial definida e na concentração de HIF-1α, e isso pode, por sua vez fornecer sugestões adequadas para as condições iniciais de cultura e tempos de incubação. Os resultados deste estudo ajudarão a lançar luz sobre as interações entre substitutos ósseos baseados em nano e sistemas biológicos, que podem servir para promover o desenvolvimento de biomateriais inovadores para uso em medicina regenerativa.
Materiais e métodos
Preparação e caracterização de partículas
HANPs a 20 nm (np20), 20 * 80 nm (np80) e partículas de HA de micro-tamanho (m-HAP) com ≥ 99,0% de pureza foram adquiridos da Nanjing Emperor Nano Material Company Ltd (Nanjing, China). O tamanho e a forma das partículas foram visualizados usando microscopia eletrônica de transmissão (TEM; FEI Tecnai G2 Spirit Bio-Twin, FEI, Hillsboro, OR, EUA) e microscopia eletrônica de varredura (SEM; LEO1530VP, Alemanha). O tamanho hidrodinâmico e o potencial zeta das partículas de HA (HAPs) foram determinados via Zetasizer Nano ZS90 e Mastersizer 3000 (Malvern Instruments, Malvern, Reino Unido).
Preparação e cultura de células
Todos os protocolos experimentais foram aprovados pelo Comitê de Ética da Universidade Médica de Nanjing. HUVECs e hWJ-MSCs foram colhidos de cordões umbilicais humanos frescos, conforme descrito anteriormente, após obter o consentimento informado por escrito dos doadores [16, 17]. Resumidamente, o cordão umbilical e a veia umbilical foram enxaguados com solução salina tamponada com fosfato (PBS) contendo 1% de penicilina e estreptomicina (PS; Hyclone, GE Healthcare Life Sciences, Pasching, Áustria). Em seguida, a veia umbilical foi preenchida com colagenase I 0,1% (Sigma, St. Louis, MO, EUA) e incubada por 15 min a 37 ° C. Após a coleta, os HUVECs foram cultivados em meio EC (ECM) (Sciencell, San Diego, CA, EUA).
Posteriormente, os vasos sanguíneos foram removidos e a geléia de Wharton foi cortada em 1 mm 2 peças e depois colocadas em 25 cm 2 frascos de cultura de tecidos (Corning Incorporated, Corning, NY, EUA). Estas células foram incubadas em L-DMEM (GIBCO Life Technology, Grand Island, NY, EUA) suplementado com 10% de soro fetal bovino (GIBCO) e 1% de PS.
As hWJ-MSCs foram avaliadas para confirmar o fenótipo usando anticorpos monoclonais para CD13, CD29, CD34, CD44, CD45, CD51 e CD105 (BD Biosciences, San Jose, CA, EUA). HUVECs foram avaliados usando o fator de von Willebrand (vWF; Shanghai ChangDao Biotech Co, Ltd., Shanghai China). HUVECs entre as passagens 3-7 e hWJ-MSCs entre as passagens 3-5 foram usados nestes experimentos.
Suspensões de partículas a 1 mg / mL em PBS foram então diluídas em L-DMEM para a concentração final. Como mostrado na Fig. 1, os HUVECs foram incubados na concentração indicada de HAPs por 18 h. O meio de cultura foi centrifugado a 15.000 rpm a 4 ° C por 15 min, e os sobrenadantes suplementados com FBS a 10% foram usados como o meio condicionado (CM) para hWJ-MSCs para realizar os seguintes experimentos. O MC consistia em meio osteogênico suplementado com fluido de indução osteogênico, que continha 10 mM de β-glicerofosfato, 50 μg / mL de ácido L-ascórbico-2-fosfato e 0,1 μM de dexametasona (Sigma-Aldrich, St Louis, MO, EUA). Além disso, 2-metoxestradiol (2-MeOE2) (Selleck Chemicals, Houston, TX, EUA) foi usado como um inibidor específico de HIF-1α. No grupo 2-MeOE2 (+), hWJ-MSCs foram cultivados com CM de HUVECs, que foram pré-tratados com 5 μM de 2-MeOE2 por 40 min antes da suplementação de HAP. PD98059 foi usado como o inibidor específico de MEK. No grupo PD98059, hWJ-MSCs foram cultivados com CM contendo 5 μM de PD98059.
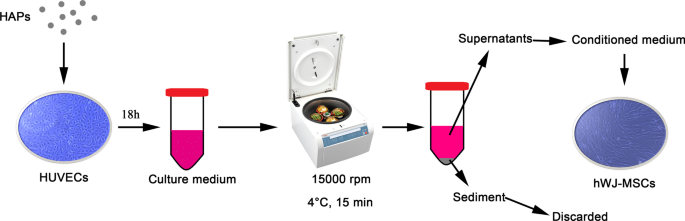
Ilustração de co-cultura indireta de HAPs / hWJ-MSCs mediada por HUVECs. Abreviações: HAPs partículas de hidroxiapatita, hWJ-MSCs células-tronco mesenquimais derivadas de gelatina de Wharton de cordão umbilical humano, HUVECs células endoteliais da veia umbilical humana
Determinando a viabilidade celular e o número
A viabilidade celular foi avaliada usando o kit de ensaio MTS (MTS; Bestbio, Beyotime Biotechnology, Shanghai, China). As hWJ-MSCs foram deixadas aderir por 24 he então cultivadas com CM por 24 e 72 h. A absorvância do formazan foi avaliada a 490 nm usando um leitor de microplacas (SpectraMax M2; Molecular Devices LLC, Sunnyvale, CA, EUA). A absorbância também foi convertida em números de células usando curvas de calibração padrão nas mesmas condições.
Reação em cadeia da polimerase quantitativa em tempo real (RT-PCR)
O reagente TRIzol (Invitrogen, Carlsbad, CA, EUA) foi usado para isolar o RNA total de células hWJ-MSCs incubadas em CM pelo tempo indicado. O DNA complementar foi transcrito a partir de 1,0 µg de RNA usando um kit PrimeScript First Strand cDNA Synthesis (TaKaRa, Tóquio, Japão) em um termociclador T3 (Mastercycler 5333; Eppendorf, Hamburgo, Alemanha). Os níveis de expressão dos genes indicados foram analisados usando o kit FastStart Universal SYBR Green Master (ROX) (Roche, Basel, Suíça) em um sistema de amplificação quantitativa em tempo real (7900HT Fast; Applied Biosystems, Foster City, CA, EUA). A expressão relativa do mRNA do gene alvo foi normalizada para GAPDH e então determinada usando o método \ (2 ^ {{- \ Delta \ Delta C_ {t}}} \). As sequências de primer para os genes alvo estão listadas na Tabela 1.
Coloração com vermelho de alizarina S (ARS) e análise quantitativa
Os hWJ-MSCs foram cultivados em placas de 12 poços com meio osteogênico por até 14 dias e, em seguida, a mineralização da matriz extracelular foi observada usando coloração ARS (Leagene, Leagene Biotechnology, Pequim, China) após o. Resumidamente, as amostras foram fixadas com álcool etílico absoluto por 15 min e, em seguida, coradas em 1% (p / v) ARS (pH, 4,2) em temperatura ambiente por 5 min. As células coradas foram lavadas duas vezes com água bidestilada e depois fotografadas. Para análise quantitativa do processo de mineralização, 300 μL de cloreto de cetilpiridínio mono-hidratado a 10% (p / v) (BOMEI, BOMEI Biotechnology, Hefei, China) foram adicionados a cada poço e as placas foram incubadas por 30 min. Um total de 90 μL de cada amostra foi transferido para uma placa de 96 poços, e a absorbância foi medida a 405 nm em triplicata.
Coloração com Fosfatase Alcalina (ALP) e Análise Quantitativa
Depois que as hWJ-MSCs foram cultivadas em placas de 12 poços com meio osteogênico por até 14 dias, a coloração de ALP foi conduzida usando um kit de desenvolvimento de cor de fosfatase alcalina BCIP / NBT (Beyotime). Resumidamente, hWJ-MSCs foram fixados em paraformaldeído a 4%. Em seguida, as amostras foram coradas em uma mistura de nitro-azul de tetrazólio e 5-bromo-4-cloro-3-indolil-fosfato por 4 he fotografadas. Para quantificar a síntese de ALP, as células foram lisadas em tampão de lise RIPA gelado (Beyotime) por 30 min. Os lisados celulares foram centrifugados a 12.000 rpm a 4 ° C por 10 min, e os sobrenadantes foram submetidos à análise quantitativa de ALP usando um kit de ensaio ALP (Beyotime). A absorvância foi medida em triplicado a 405 nm e convertida em atividade de ALP usando uma curva padrão.
Ensaio de imunoabsorção enzimática (ELISA)
HUVECs foram semeados em 2 × 10 5 células / poço. CM foi coletado de HUVECs cultivados com HAPs por 18 h (arquivo adicional 1), e o sobrenadante foi submetido à análise de ELISA (Fig. 1). Um kit HIF-1α ELISA humano (Anhui Joyee Biotechnics, Anhui, China) foi usado de acordo com as instruções do fabricante.
Imunofluorescência
HUVECs foram semeados em lâminas em placas de 12 poços (7,6 × 10 4 células / poço). Após a exposição a CM por 18 h, as células foram fixadas em paraformaldeído 4% (Biosharp, Pequim, China) e permeabilizadas com Triton X ‐ 100 0,1% (Beyotime) em PBS antes da incubação com anticorpo anti-HIF-1α 1% [EPR16897 ] (ab179483, Abcam, Reino Unido) durante a noite a 4 ℃. Posteriormente, as células foram incubadas com 1% CoraLite594 — IgG anti-coelho de cabra conjugado (H + L) (Proteintech, EUA) no escuro por 1 h. Em seguida, os núcleos foram tingidos com DAPI (Beyotime, Shanghai, China), que foi adicionado às células e reagiu por 30 s. As amostras foram examinadas usando um microscópio confocal a laser (Olympus, Japão). A intensidade de fluorescência foi quantificada usando o software de análise ImageJ v.1.4 (Bethesda, MD, EUA).
Análise de Western Blot
hWJ-MSCs foram incubados em CM por 24 h (quinase regulada por sinal extracelular (ERK) 1/2, p-ERK1 / 2) ou em meio osteogênico por 7 d (fator de transcrição relacionado ao runt (RUNX) -2, tipo 1 colágeno / colágeno 1 (COL I)). Em seguida, as células foram lisadas em tampão de lise RIPA por 30 min. Os lisados celulares foram centrifugados e os sobrenadantes foram armazenados a -20 ° C para análise. Após SDS-PAGE 12%, as proteínas foram transferidas para uma membrana de difluoreto de polivinilideno (PVDF). Os anticorpos primários usados foram anti-p-ERK1 / 2, anti-ERK1 / 2, anti-RUNX-2, anti-COL I e GAPDH (1:1.000, anticorpos policlonais de coelho; Cell Signaling Technology, Boston, MA, EUA ) Após a remoção dos anticorpos não ligados, a membrana foi incubada com anticorpos secundários por 1 h. O sinal nas membranas foi detectado usando um sistema de imagem em gel de quimioluminescência (LAS4000M; GE Healthcare Biosciences AB, Uppsala, Suécia). A proporção de p-ERK para ERK e RUNX-2 / COL I para GAPDH foi quantificada usando o software de análise ImageJ v.1.4 (Bethesda).
Avaliação da apoptose celular
hWJ-MSCs foram semeados a uma densidade de 10 5 células por poço em placas de 6 poços. As células aderentes foram tratadas com as concentrações indicadas de HIF-1α durante o tempo indicado. As células foram então coletadas e marcadas com FITC-Anexina V e PI (Fcmacs, Nanjing, China) por 15 min no escuro. Todas as amostras foram testadas usando um citômetro de fluxo FACScan (BD Bioscience, San Jose, CA, EUA). Os dados foram analisados usando FlowJo v10 (BD Biosciences).
Modelo de linhagem celular de dois estágios
Em consideração ao enorme potencial dos HAPs e à dificuldade em analisar o sistema de co-cultura, foi necessário um modelo matemático que pudesse fornecer uma análise quantitativa e uma previsão confiável para entender o papel do HIF-1α na diferenciação osteogênica de hWJ- MSCs.
As hWJ-MSCs foram cultivadas com 0, 300, 500, 1000, 1500, 2000, 3000 e 4000 pg / mL de HIF-1α, bem como fluido de indução osteogênico. Após ajustar os dados a essas concentrações, usamos as concentrações de HIF-1α (produzido por HUVECs) nos grupos de controle, m-HAP, np80 e np20 (240, 300, 325 e 375 pg / mL, respectivamente) para teste as equações de ajuste usando o MATLAB (MathWorks, Natick, MA, EUA). Para simplificar o modelo, consideramos seus padrões de crescimento semelhantes nas diferentes concentrações de HIF-1α como sendo idênticos. O grau de diferenciação médio foi empregado para ajustar a equação grau de diferenciação-tempo. A taxa de proliferação, taxa de apoptose e graus de diferenciação osteogênica das hWJ-MSCs nos diferentes grupos foram detectados em momentos definidos.
Um modelo simplificado de linhagem celular de dois estágios, que era semelhante a um modelo de linhagem celular de múltiplos estágios [18, 19], foi estabelecido de acordo com os dados experimentais. C 0 e C 1 representam o número de células das hWJ-MSCs e células terminais, respectivamente. C 0 e C 1 são regidos por:
$$ \ left \ {\ begin {reuniu} \ frac {{{\ text {d}} C_ {0}}} {{{\ text {d}} t}} =\ left [{\ frac {{K - C_ {0} - C_ {1}}} {K} p - (p - 1)} \ direita] \ upsilon_ {0} C_ {0} \ hfill \\ \ frac {{{\ text {d}} C_ {1}}} {{{\ text {d}} t}} =\ left ({2 - \ frac {{K - C_ {0} - C_ {1}}} {K} p - p} \ direita) \ upsilon_ {0} C_ {0} - AC_ {1} \ hfill \\ \ end {recolhido} \ certo. $$
Aqui, p , afetado por HIF-1α e tempo, representa a probabilidade de replicação dos hWJ-MSCs. Correspondentemente, d =1 - p é a taxa de diferenciação que pode ser obtida ajustando os dados estimados e experimentais do número de células. O parâmetro v 0 quantifica a rapidez com que as células se dividem em cada estágio de linhagem (em particular, v =Ln2 / c , onde c é a duração de um ciclo celular). A taxa de apoptose das células terminais é simbolizada por A . Para simplificar, negligenciamos que a taxa de apoptose irá variar ligeiramente ao longo do tempo e, portanto, A =4,5% é uma constante. K denota a capacidade ambiental porque as células não poderiam sofrer proliferação ilimitada [20]. HIF-1α aumenta a taxa de diferenciação dos hWJ-MSCs, levando a uma taxa de diferenciação modelada por:
$$ \ begin {alinhados} d &=\ frac {{d_ {0}}} {{1 + (r * H) ^ {m}}} \\ p &=1 - d \\ \ end {alinhados} $$
Aqui, H representa a concentração de HIF-1α; d 0 denota a taxa de diferenciação em 0 pg / mL HIF-1α; r representa a intensidade de regulação (aqui, representando a intensidade de regulação de HIF-1α em MSCs); e m corresponde ao coeficiente de Hill [21], scilicet a relação entre a taxa de diferenciação de MSC e a concentração de HIF-1α.
Análise estatística
Todos os dados que atendem aos requisitos de normalidade e homocedasticidade são expressos como a média ± desvio padrão (DP) de três ou mais experimentos independentes. O software SPSS 24.0 (SPSS Inc., Chicago, EUA) foi usado para realizar as análises estatísticas por meio de ANOVA de uma via ou ANONA de duas vias. A P valor <0,05 foi considerado estatisticamente significativo. A análise estatística é apresentada usando GraphPad Prism 5 (GraphPad Software, La Jolla, CA).
Resultados
Caracterização de HAPs
Como mostrado na Fig. 2, os HAPs foram preparados com tamanho e forma particulares. Os diâmetros do np20 com formato quase esférico eram de 20 nm em média, e o np80 era semelhante a uma haste com comprimento médio de 80 nm e largura de 20 nm. Os m-HAPs também eram quase esféricos em forma e aproximadamente 12 μm de diâmetro. Todas as partículas tinham uma carga superficial negativa em L-DMEM. Foi sugerido que os valores negativos do potencial zeta têm um efeito favorável significativo na fixação e proliferação de células ósseas, bem como na ligação direta do osso e na formação de novo osso [22, 23]. As partículas, que foram observadas em L-DMEM, têm tendência a se agregar em sistemas aquosos. Seu tamanho hidrodinâmico também foi testado, o que também pode ser um fator importante que afeta seus comportamentos biológicos.

Caracterização de HAPs. Micrografias TEM de a np20 e b np80 e micrografia SEM de c m-HAP. d Caracterização de HAPs ( n =6). Abreviações: TEM microscopia eletrônica de transmissão, SEM microscopia eletrônica de varredura, HA hidroxiapatita, m-HAP partículas de HAP de tamanho micro
Toxicidade indireta de HAPs para hWJ-MSCs
Para avaliar a toxicidade indireta de HAPs em hWJ-MSCs, a viabilidade celular foi medida por meio de ensaios MTS. CM com HANPs de 50 µg / mL pode estimular significativamente a viabilidade de hWJ-MSC após 24 he 72 he especialmente em 24 h. No entanto, o CM com HANPs de 100 µg / mL diminuiu a viabilidade celular em 15–20% em comparação com o controle após 72 h. Além disso, CM com 25 µg / mL np20, mas não np80, estimulou a viabilidade celular após 24 h. Esses fenômenos confirmaram que 50 µg / mL de HANPs eram uma concentração subcitotóxica que foi usada em todos os experimentos subsequentes (Fig. 3).
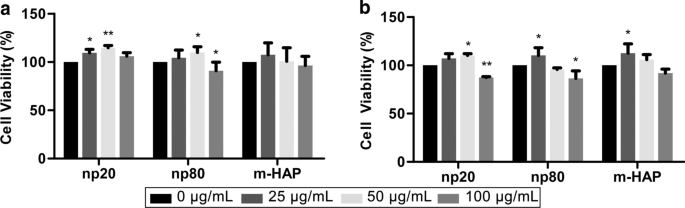
Toxicidade indireta de HAPs para hWJ-MSCs. A viabilidade de hWJ-MSCs cultivados indiretamente com HAPs foi medida para a 24 e b 72 h. * P <0,05; ** P <0,01 versus o controle. O grupo controle consistiu em células incubadas em CM sem tratamento com HAPs, e a viabilidade celular foi normalizada como uma porcentagem do controle. Abreviações: HAPs partículas de hidroxiapatita, m-HAP partículas de HAP de tamanho micro, hWJ-MSCs Células-tronco mesenquimais derivadas de geléia de Wharton do cordão umbilical humano
Efeito osteoindutivo indireto de HAPs em hWJ-MSCs
Para identificar os efeitos osteoindutivos indiretos de HANPs em hWJ-MSCs, a expressão de genes osteogênicos foi avaliada por análise quantitativa de RT-PCR. O nível de transcrição para o fator de transcrição relacionado ao runt 2 (RUNX-2) nos grupos HANP, especialmente np20, exibiu um aumento notável do Dia 7 ao Dia 21 (Fig. 4a). A expressão gênica do colágeno tipo I (Col I) no grupo np20 demonstrou um aumento do dia 7 ao dia 21, enquanto o grupo np80 demonstrou um aumento sustentado do dia 7 ao dia 14 (Fig. 4b). O mRNA da osteocalcina (OCN) foi claramente regulado positivamente nos grupos HANP no Dia 14, indicando uma taxa acelerada de osteogênese (Fig. 4c). Os níveis de mRNA de fosfatase alcalina (ALP) obviamente aumentaram nos grupos HANP (Fig. 4d), indicando a diferenciação osteogênica de hWJ-MSCs. Porém, o grupo m-HAP apresentou alterações limitadas nos níveis desses três genes osteogênicos em relação ao grupo controle (fig. 4). Além disso, a expressão dos marcadores de pluripotência, NANOG, OCT3 / 4 e SOX2, diminuiu nos grupos HANPs em comparação com o controle (Fig. 4e-g), implicando que os hWJ-MSCs nos grupos HANPs se diferenciaram, especialmente em o grupo np20. Resultados semelhantes foram obtidos por análise de Western blot (Fig. 5c, d), indicando que o grupo np 20 pode aumentar indiretamente a expressão de RUNX-2 e COL I em hWJ-MSCs.
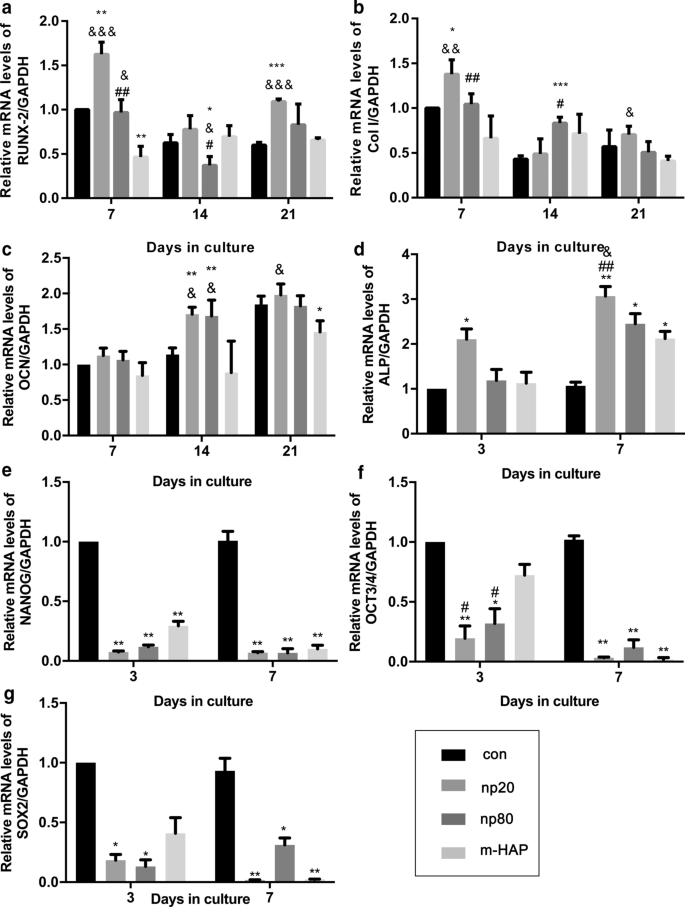
Efeitos indiretos dos HAPs na expressão de genes relacionados à diferenciação osteogênica. a RUNX-2, b Col I, c OCN, d ALP, e NANOG, f OCT3 / 4 e g Níveis do gene SOX2 em hWJ-MSCs cultivadas com CM pelo tempo indicado. * P <0,05; ** P <0,01; *** P <0,001 versus grupo controle; e P <0,05; && P <0,01; &&& P <0,001 versus o grupo m-HAP; # P <0,05; ## P <0,01 versus o grupo np20. As células incubadas em meio osteogênico sem tratamento com HAPs foram utilizadas como grupo de controle. Abreviações: HAPs partículas de hidroxiapatita, m-HAP partículas de HAP de tamanho micro, hWJ-MSCs células-tronco mesenquimais derivadas de gelatina de Wharton de cordão umbilical humano, RUNX-2 fator de transcrição 2 relacionado ao runt, Col I colágeno tipo I, OCN osteocalcina, ALP fosfatase alcalina, SOX 2 HMG-box 2 relacionado com SRY
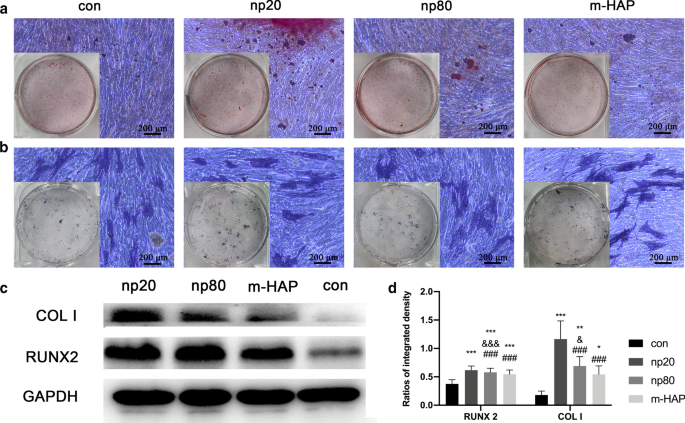
Efeito indireto dos HAPs na deposição de cálcio extracelular e na atividade de ALP. As hWJ-MSCs foram incubadas em meio osteogênico por 14 dias. a A deposição de cálcio extracelular foi então visualizada por meio de coloração ARS; b A atividade de ALP das hWJ-MSCs foi avaliada por meio de coloração de ALP, barras de escala:200 μm. c A análise de Western blot indicou a expressão de RUNX-2 e COL I de hWJ-MSCs em meio osteogênico no dia 7. d Medidas densitométricas de RUNX-2 e COL I da parte ( c ) As células incubadas em meio osteogênico sem tratamento com HAPs foram utilizadas como grupo de controle. * P <0,05; ** P <0,01; *** P <0,001 versus grupo controle; e P <0,05; &&& P <0,001 versus o grupo m-HAP; ### P <0,001 versus o grupo np20. Abreviações: HAPs partículas de hidroxiapatita, m-HAP partículas de HAP de tamanho micro, hWJ-MSCs células-tronco mesenquimais derivadas de gelatina de Wharton do cordão umbilical humano, ALP fosfatase alcalina
Para observar visualmente o efeito osteoindutor indireto de HANPs em hWJ-MSCs, as células foram incubadas com o meio osteogênico indicado por 14 dias, seguido de coloração ARS e ALP. Como mostrado na Fig. 5a, b, um número aumentado de nódulos mineralizados e maior atividade ALP das hWJ-MSCs foram observados nos grupos HANP em comparação com os grupos m-HAP e controle. Além disso, o m-HAP, semelhante ao controle, demonstrou efeitos limitados na diferenciação osteogênica das hWJ-MSCs.
Sinalização ERK1 / 2 ativada por HAPs em hWJ-MSCs indiretamente co-cultivados com HUVECs
Para investigar os efeitos sobre a função parácrina de HUVECs por HAPs, ensaios de imunofluorescência e ELISA foram usados para identificar a possível proteína promovendo a diferenciação osteogênica das hWJ-MSCs. Conforme mostrado na Fig. 6a-c, a produção intracelular e extracelular de HIF-1α foi significativamente facilitada por HANPs (especialmente np20), embora houvesse um efeito limitado de m-HAP em sua produção.
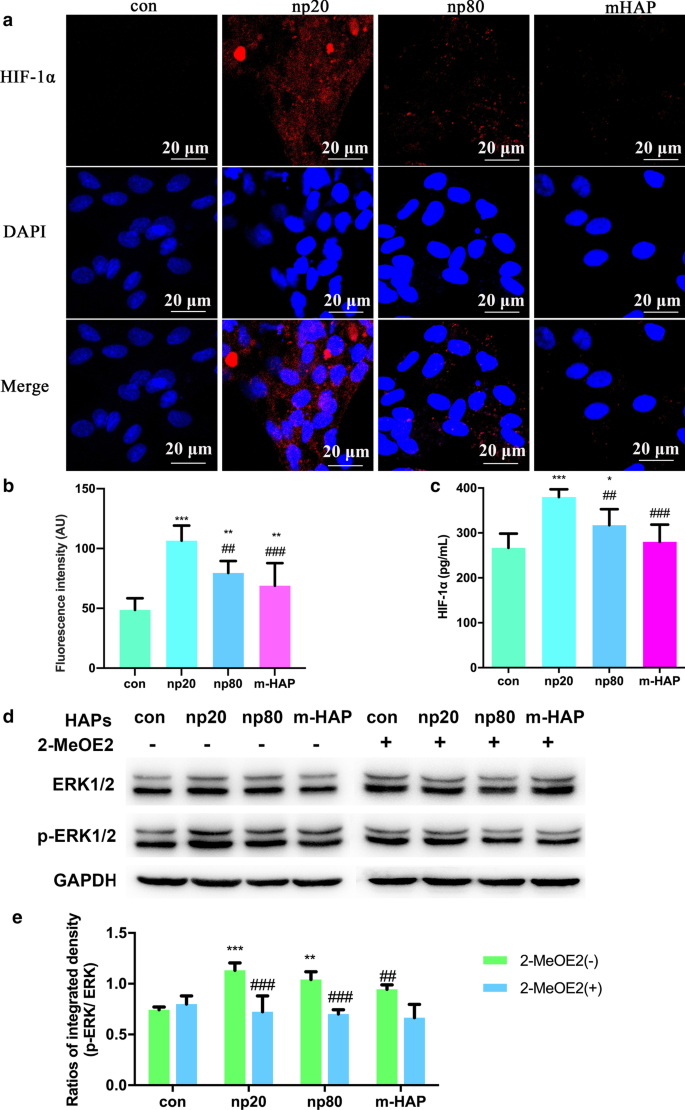
Os HAPs ativaram a sinalização de ERK1 / 2 em hWJ-MSCs indiretamente co-cultivados com HUVECs. a Imunofluorescência de HIF-1α realizada em HUVECs tratados com / sem HAPs por 18 h. b A intensidade de fluorescência de HIF-1α da parte ( a ) c A concentração extracelular de HIF-1α no meio de cultura de HUVECs tratados com / sem HAPs por 18 h foi medida por meio de ELISA. Barras de escala:20 μm. * P <0,05; ** P <0,01; *** P <0,001 versus controle; ## P <0,01; ### P <0,001 versus o grupo np20. As células sem tratamento com HAPs foram utilizadas como grupo de controle. hWJ-MSCs foram tratados com CM por 24 h. d Análise de Western blot indicando ativação de quinases-chave nas vias ERK1 / 2. e Medições densitométricas de p-ERK1 / 2 da parte ( b ) ** P <0,01; *** P <0,001 versus controle; ## P <0,01, ### P <0,001 versus grupo 2-MeOE2 (-). As células incubadas em CM sem tratamento com HAP foram utilizadas como grupo de controle. Abreviações: HAPs partículas de hidroxiapatita, m-HAP partículas de HAP de tamanho micro, hWJ-MSCs células-tronco mesenquimais derivadas de gelatina de Wharton de cordão umbilical humano, HUVECs células endoteliais da veia umbilical humana, ERK quinase regulada por sinal extracelular, HIF-1α fator 1α induzível por hipóxia
Para compreender com mais precisão a via de sinalização de diferenciação dos hWJ-MSCs ativados por HIF-1α, examinamos os principais reguladores da via ERK1 / 2 por meio de análise de Western blot. Como mostrado na Fig. 6d, e, enquanto os níveis de proteína de ERK1 / 2 total permaneceram inalterados, os níveis de p-ERK1 / 2 foram nitidamente aumentados nas hWJ-MSCs cultivadas com HANPs, e isso foi especialmente verdadeiro no grupo np20. No entanto, m-HAP teve pouco efeito nos níveis de p-ERK1 / 2 nos hWJ-MSCs, semelhante ao seu efeito na produção de HIF-1α em HUVECs. É importante ressaltar que os níveis aumentados de p-ERK1 / 2 nos hWJ-MSCs ativados por HIF-1α podem ser bloqueados por 2-MeOE2, um inibidor específico de HIF-1α, indicando que HIF-1α funcionou a montante da via de sinalização ERK1 / 2 em hWJ-MSCs.
Diferenciação Osteogênica promovida por HIF-1α de hWJ-MSCs por meio da via ERK1 / 2
Para determinar se o HIF-1α era necessário para a estimulação observada da diferenciação osteogênica de hWJ-MSC, um inibidor específico de HIF-1α (2-MeOE2) foi aplicado a essas culturas de células. Como mostrado na Fig. 7, a deposição da matriz mineralizada e a atividade ALP do grupo tratado com 2-MeOE2 (+) - hWJ-MSCs cultivados em meio osteogênico foram enfraquecidas, indicando que o HIF-1α foi indispensável para a diferenciação osteogênica do hWJ- MSCs. Com base nesses resultados, exploramos ainda o papel da via ERK1 / 2 na diferenciação osteogênica das hWJ-MSCs ativadas por HIF-1α. A deposição da matriz mineralizada e a atividade ALP nas hWJ-MSCs cultivadas com meio osteogênico foram suprimidas após a administração de PD98059, um inibidor específico de MEK.
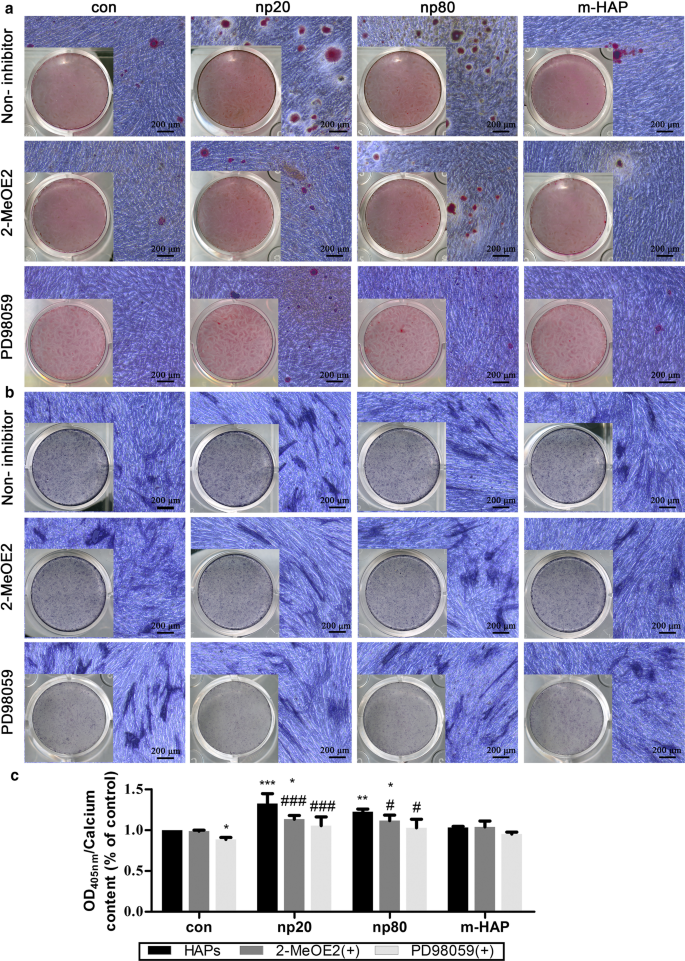
O HIF-1α promoveu a diferenciação osteogênica de hWJ-MSCs através da via ERK1 / 2. As hWJ-MSCs foram incubadas em meio osteogênico com ou sem PD98059 por 14 dias. a A deposição de cálcio extracelular foi visualizada por meio de coloração ARS. b A atividade de ALP de hWJ-MSCs foi avaliada por meio de coloração de ALP. Barras de escala:200 μm. c Análise quantitativa da matriz extracelular de cálcio. * P <0,05; ** P <0,01; *** P <0,001 versus controle; # P <0,05; ### P <0,001 versus o grupo np20. As células incubadas em meio osteogênico sem HAPs e tratamento com PD98059 foram usadas como grupo de controle. Abbreviations:HAPs hydroxyapatite particles, m-HAP micro-sized HAP particles, hWJ-MSCs human umbilical cord Wharton’s jelly-derived mesenchymal stem cells, HUVECs human umbilical vein endothelial cells, ERK extracellular signal-regulated kinase, HIF-1α hypoxia-inducible factor 1α, ALP alkaline phosphatase
Two-Stage Cell-Lineage Models
To quantitatively reveal the intrinsic connection between the concentration of HIF and osteogenic differentiation of hWJ-MSCs, 0–4000 pg/mL of HIF-1α was used to treat eight groups of hWJ-MSCs. A two-stage cell-lineage mathematical model was used to analyze the proliferation, apoptosis, and osteogenic differentiation rates of these hWJ-MSCs treated with different concentrations of HIF-1α. As shown in Fig. 8a, fitting data were employed to obtain the simulated formula (\(\frac{d}{{d_{0} }} =\frac{1}{{0.14H^{2} - 0.43H + 1}}\)) and curve (blue curve), showing that the differentiation rate first increased and then decreased with the increase in HIF concentration. More specifically, the differentiation rate reached a peak at 1500 pg/mL HIF-1α.

Two-stage cell-lineage models. hWJ-MSCs were incubated in a defined concentration of HIF-1α for the indicated times. a Relative differentiation rates of hWJ-MSCs at different concentrations of HIF-1α, b relative ALP activity (differentiation degrees of hWJ-MSCs) and relative osteoblast cells on different culture days. c Three-dimensional surface of differentiation degree evolving with time and HIF-1α. Cell number with an initial seeding density of 1500 cells per well (in a 96-well plate), as well as d 0, e 375, and f 1500 pg/mL HIF-1α. Abbreviations:hWJ-MSCs human umbilical cord Wharton’s jelly-derived mesenchymal stem cells, HIF-1α hypoxia-inducible factor 1α, ALP alkaline phosphatase
According to our study, the concentrations of HIF-1α produced by HUVECs in the control, m-HAP, np80, and np20 groups were 240, 300, 325, and 375 pg/mL, respectively. The black square represents the differentiation rate promoted by 240, 300, 325, and 375 pg/mL HIF-1α, and this matches well with the simulated curve. In addition, the differentiation degrees of the hWJ-MSCs (relative ALP activity) treated with different concentrations of HIF-1α increased similarly increased with time. Therefore, in order to simplify the model, we consider their growth patterns to be similar or even identical. We found that the increase in ALP activity from Day 0 to Day 7 was proportional to the number of osteoblasts, and osteoblast cells reached their peak at the platform period. Therefore, after fitting the ALP activity with the relative osteoblasts cell number curve (osteoblasts cell number / maximum osteoblasts cell number), the maximum of ALP activity was predicted and relative ALP activity (ALP activity/ maximum of ALP activity), representing the differentiation degree, was acquired (Fig. 8b). Combining these two studies, the three-dimensional surface of the differentiation degree evolving with time and HIF-1α was obtained (Fig. 8c).
In order to estimate the optimal culture time, it was necessary to simulate cell numbers. After elucidating the differentiation rate under different concentrations of HIF-1α, we simulated the size of each population. The simulation utilized an initial seeding density of 1500 cells per well (in a 96-well plate) using different concentrations of HIF-1α. As shown in Fig. 8d–f, the experimental data (black square) match well with the simulated total cell numbers (blue curve), which is a sum of the number of hWJ-MSCs and osteoblasts, and this supports the ability of this model to predict the number of hWJ-MSCs and osteoblasts at any time point. Moreover, the osteoblast cell number approached the platform period at 21, 18, and 15 days with 0, 375, and 1500 pg/mL HIF-1α, respectively. This model provides the optimum culture time for guiding tissue engineering, and it also provides direct evidence that HIF-1α accelerates the osteogenic differentiation of hWJ-MSCs.
Discussão
With recent advances in nanobiomaterials, nano-based artificial bone substitutes have been an area of intense investigation. The accumulating evidence suggests that there are complex interactions between cells and nanobiomaterials due to their capacity to penetrate cell membranes and increase internal retention times [24, 25]. A previous study revealed that collagen/alginate nanofilms can adsorb onto the MSC membrane to activate intracellular signaling cascades and promote osteogenic differentiation [26]. Elegant experiments by Wu and his colleagues clearly demonstrated that TiO2 nanotubes can improve vascularization and osteogenic differentiation by facilitating paracrine effects and cell junctions via EC-MSC interactions [27]. For the purpose of developing excellent candidates for bone tissue engineering, it is necessary to clarify the direct crosstalk between nano-based bone substitutes and cells implicated in bone repair as well as their indirect interactions. However, our current understanding of this is still limited. In the present study, we utilized an indirect co-culture model to further elucidate the biological effects of HANPs on MSCs in regard to the indirect interactions mediated by ECs.
Cytotoxicity is a primary issue for assessing the biocompatibility of any nanobiomaterial. Although our previous study found that HANPs did not directly influence the viability or apoptosis of hWJ-MSCs, they may still exert different impacts via the mediation of other cells [28]. Thus, it was necessary to evaluate the cytotoxic effects of HANPs on hWJ-MSCs mediated by HUVECs. Interestingly, after incubation in CM for 24 h and 72 h, hWJ-MSC viability was maintained and even elevated in the 0–50 µg/mL HANP groups, especially in the np20 group, indicating the existence of effector molecules in the CM. When the concentration of HANPs reached 100 µg/mL, they became cytotoxic to the hWJ-MSCs. However, 0–100 µg/mL m-HAP had no influence on hWJ-MSC viability (Fig. 3). Jiang et al. have shown that engineered nanoparticles of a particular size can have distinct endocytic routes and kinetics associated with altered downstream signaling involved in regulating target cell functions [29]. In our previous study, we showed that np20 and np80 were endocytosed by HUVECs, and this was followed by morphologic changes and the appearance of large vacuoles, indicating the activated state of the HUVECs. Additionally, np20, with their faster uptake speed and increased accumulation, might result in a stronger activation of HUVECs, possibly resulting in increased hWJ-MSC viability via paracrine signaling. Conversely, few m-HAPs can be endocytosed by HUVECs, and this might account for their limited influence on the metabolism of hWJ-MSCs [9].
To further explore the potential osteoinductive effect of activated HUVECs, a subcytotoxic dose of 50 µg/mL HAPs was used in subsequent studies. The CM collected from the activated HUVECs promoted extracellular calcium deposition, ALP activity, and osteogenic proteins expression in hWJ-MSCs, as well as the mRNA expression of osteogenic genes (Figs. 4, 5). Runx2, an essential transcription factor involved in specifying the osteoblast lineage [30], showed a substantial enhancement in the np20 group, indicating a strong osteoinductive effect on hWJ-MSCs (Fig. 4a and Fig. 5c, d). The np20 group demonstrated a 1.5-fold improvement in COLI expression at Day 7 (Figs. 4b, 5c, d) and a double increase at Day 14, which implied the presence of additional differentiated osteoblasts in the HANP-treated groups (Fig. 4b) [30]. OCN is a mature stage bone marker [31], and this gene showed a significant increase in the HANP groups at Day 14 (Fig. 4c), indicating that np20 and np80 can accelerate bone maturation compared to m-HAP. ALP is an early marker of osteoblast differentiation, and it obviously increased with culture time in each group, especially the np20 group, revealing that additional transformation occurred from MSCs to osteoblasts (Fig. 4d). Pluripotency markers, NANOG, OCT3/4, and SOX2 imply the capacity for differentiation [32]. As shown in Fig. 4e–g, the decreased expression in the genes of the HANP groups implied that most of the hWJ-MSCs in HAP groups had transformed into osteoblasts.
Our data demonstrated that the endocytosis of HANPs by HUVECs was associated with an improved osteogenic differentiation of hWJ-MSCs. However, the cause of this outcome is currently unclear. In terms of the paracrine function of HUVECs, we focused on soluble differentiation-inducing proteins in the supernatant of activated HUVECs. HIF-1α signaling is essential in coupling ossification and angiogenesis during bone regeneration [33, 34]. Heikal et al. reported that injured ECs secrete more HIF-1α even under normoxia conditions [35]. It has also been shown that exposure to HANPs inhibits the angiogenic ability of HUVECs [9]. Thus, we measured the concentrations of HIF-1α in the CM, and the results showed that the HIF-1α content increased in the HANP treatment groups compared to the m-HAP and control groups (Fig. 6a). To identify the role of HIF-1α in the osteogenic differentiation of hWJ-MSCs, we used 2-MeOE2, which is a specific HIF-1α inhibitor, was used. The decreased concentration of HIF-1α paralleled the impaired mineralized matrix deposition and ALP activity in these hWJ-MSCs, indicating that HANPs can promote the HIF-1α production of HUVECs to facilitate the osteogenesis of hWJ-MSCs (Fig. 7).
To properly apply HANPs for use in bone tissue engineering, it is necessary to gain further insights into the mechanisms by which HANPs promote the osteogenic differentiation of hWJ-MSCs mediated by HUVECs. The ERK1/2 pathway is downstream of HIF-1α [36] and is fundamental to the differentiation of MSCs [37]. In this work, the concentrations of HIF-1α in the CM coincide well with the p-ERK1/2 levels in the hWJ-MSCs (Fig. 6b, c). When 2-MeOE2 was applied, the p-ERK1/2 expression in the hWJ-MSCs failed to be activated, indicating that HIF-1α functioned upstream of ERK1/2 signaling. To directly address the role of ERK1/2 signaling in the osteogenic differentiation of hWJ-MSCs, PD98059, a specific MEK inhibitor, was used. The suppression of ERK1/2 signaling resulted in the lowest osteogenic differentiation of hWJ-MSCs. One possible reason for this occurrence is that the ERK1/2 pathway plays a key role in both HIF-1α signaling and in the apoptosis and proliferation signaling pathways, which could be responsible for the observed changes in osteogenic differentiation in these cells [38, 39]. Additionally, this could also be related to the presence of vascular endothelial growth factor (VEGF). VEGF is one of the downstream effectors of HIF-1α signaling [33], and it can also promote the osteogenic differentiation of MSCs via activation of the ERK1/2 pathway [37]. Our previous study found that np20 induced the production of VEGF in HUVECs [9]; therefore, it is possible that the suppression of the ERK1/2 pathway may result in inhibition of VEGF, which would lead to the decreased osteogenic differentiation of hWJ-MSCs. According to the available experimental results, we can summarize as follows. HANPs are able to more optimally process better direct [5] and indirect osteoinductive effects than m-HAPs. Compared to autogenous bone grafts and bone allografts, there is an extensive source of HANP and without secondary damage and potential immunogenicity. However, compared to m-HAPs, HANPs can suppress the angiogenic ability of HUVECs [9] and exhibit slight cytotoxicity in both a time- and dose-dependent fashion.
Recently, growing evidence has demonstrated the importance of HIF-1α in the bone regeneration. However, few studies have been able to quantitatively predict the MSC differentiation rate under specific initial conditions, such as the HIF-1α concentration. Taking cell proliferation, apoptosis, and osteogenic differentiation into account, we present a mathematical model that combines a two-stage cell lineage with HIF-1α that is highly correlated with our experimental data. By fitting the differentiation rate of hWJ-MSCs in 0–4000 pg/mL HIF-1α, we acquired the equations for describing the differentiation rate, HIF-1α concentration, and time. As shown in Fig. 8d, this model can depict the cell number map under different HIF-1α concentrations, so that it is possible to explore the intrinsic dynamics of the two-stage system [40]. Additionally, this model mathematically validates the effect of HIF-1α on the osteogenic differentiation of hWJ-MSCs. Moreover, based on a multi-stage cell-lineage model and logistic model, our model is sufficiently stable to enable long-term predictions without falling into the trap of population unlimited explosion [41].
By using the existing experimental data, both the cell number and differentiation rate can be predicted with a defined initial cell seeding density and HIF-1α concentration. As such, the optimum incubation time is also obtained. Consequently, we can predict the optimum concentration of HIF-1α and determine the most optimal time for osteogenesis, which is important for efficient tissue engineering. A two-stage cell-lineage model is applicable for predicting the proliferation and differentiation of stem cells, which have two cell lineages. On this basis, the model founded on the initial conditions and existing experimental data can be established to identify the optimum culture conditions in vitro, which will assist in optimizing bone repair in vivo.
Conclusion
In this study, we explored the specific biological effects of HANPs on hWJ-MSCs mediated by HUVECs. Compared to m-HAPs, both np20 and np80 showed slight cytotoxicity in both a time- and dose-dependent fashion. Importantly, the size of the HANPs appeared to have no significant impact on this cytotoxicity. Our data also showed that HANPs, especially np20, were capable of facilitating HUVECs to secrete increased levels of HIF-1α, which directly correlated with the enhanced osteogenic differentiation of hWJ-MSCs via the activation of the ERK1/2 pathway (Fig. 9). More remarkably, the results from the two-stage cell-lineage model suggested that HIF-1α exerted a dose-dependent stimulatory effect on the osteogenic differentiation rate of hWJ-MSCs. Additionally, the optimum concentration of HIF-1α and incubation time were estimated based on the initial conditions using an in vitro model, which could be invaluable in the future for tissue engineering applications. Collectively, these observations provide evidence that HANPs may improve bone regeneration by modulating cell–cell interactions.

A schematic illustration of the possible mechanisms
Disponibilidade de dados e materiais
The datasets used and analyzed during the current study are available from the corresponding author on reasonable request.
Abreviações
- HA:
-
Hydroxyapatite
- HANPs:
-
HA nanoparticles
- hWJ-MSCs:
-
Human umbilical cord Wharton’s jelly-derived mesenchymal stem cells
- HUVECs:
-
Human umbilical vein endothelial cell
- m-HAP:
-
Micro-sized HAP particles
- PBS:
-
Phosphate-buffered saline
- ECM:
-
EC medium
- 2-MeOE2:
-
2-Methoxyestradiol
- ELISA:
-
Enzyme-linked immunosorbent assay
- RUNX-2:
-
Runt-related transcription factor 2
- Col I:
-
Type I collagen
- OCN:
-
Osteocalcin
- ALP:
-
Alkaline phosphatase
- SOX 2:
-
SRY-related HMG-box 2
- ERK:
-
Extracellular signal-related kinases
- VEGF:
-
Vascular endothelial growth factor
Longitudinal Zeólita-Óxido de Ferro Biossensor de Capacitância Depositada para Interleucina-3 na Detecção de Sepse
Simulação de dinâmica molecular no mecanismo de corte no processo de usinagem híbrida de silício monocristalino
Nanomateriais
- Baterias para fins especiais
- Célula solar
- Análise de Actina e Organização de Adesão Focal em Células U2OS em Nanoestruturas Poliméricas
- Nanotécnicas Inativam células-tronco cancerosas
- Estudo in vitro da influência das nanopartículas de Au nas linhas celulares HT29 e SPEV
- Efeitos de interação na montagem de nanopartículas magnéticas
- Células endoteliais de segmentação com nanopartículas de GaN / Fe multifuncionais
- Efeitos sinérgicos de nanopartículas de Ag / BiV1-xMoxO4 com atividade fotocatalítica aprimorada
- Promoção do crescimento celular SH-SY5Y por nanopartículas de ouro modificadas com 6-mercaptopurina e um peptídeo penetrador de neurônio
- Nanocompósitos à base de óxido de grafeno decorados com nanopartículas de prata como agente antibacteriano