


Thiacalix [4] arenes Remova os efeitos inibitórios dos cátions Zn na atividade ATPase da miosina
Resumo
Numerosas anomalias reprodutivas femininas são causadas por distúrbios do músculo liso uterino (miométrio). Os metais pesados têm um efeito adverso na contratilidade do músculo liso uterino. Embora o zinco seja um elemento biogênico essencial para a maioria dos organismos, altas doses desse elemento são tóxicas. O estudo de 0,5−5 mM Zn 2+ efeito sobre a atividade da miosina S1 ATPase do útero descobriu que 5 mM de Zn 2+ os cátions têm o efeito inibitório mais pronunciado. O cálculo dos parâmetros cinéticos ( K m e V máximo , ATP ) revelou que a velocidade máxima aparente do ATP de hidrólise catalisada pela miosina na presença de 5 mM de Zn 2+ diminuiu 1,6 vezes. O valor de К m para hidrólise de ATP pela miosina S1 na presença de Zn 2+ não muda estatisticamente, embora tenda a diminuir. Foi determinado que a atividade da miosina S1 ATPase uterina não depende da concentração de Mg 2+ na presença de 5 mM de Zn 2+ . Além disso, foi demonstrado que tetrahidroxitiacalix [4] areno-tetrassulfosfonato (C-798) e tetrahidroxitiacalix [4] areno-tetrafosfonato (C-800) restaurou a atividade da miosina S1 ATPase ao nível de controle na presença de 5 mM de Zn 2+ . Um dos mecanismos mais prováveis de restaurar a ação protetora desses tiacalix [4] arenos é baseado em sua capacidade de quelar cátions de metais pesados do meio de incubação. O docking molecular de C-798 e C-800 na região S1 da miosina mostrou que esses tiacalix [4] arenos poderiam interagir com a ligação do cátion Zn por resíduos de aminoácidos da miosina próximos ao sítio ativo da ATPase. Portanto, thiacalix [4] arenos pode enfraquecer a interação entre este cátion e miosina S1. Especulou-se que os resultados obtidos poderiam ser utilizados em pesquisas futuras com o objetivo de utilizar este tiacalix [4] arenos como compostos farmacológicos no caso de intoxicações por altas concentrações de zinco.
Histórico
O problema da poluição do meio ambiente com metais pesados e a busca de formas de reduzir seu impacto sobre os objetos vivos é relevante [1, 2].
O zinco é um elemento biogênico essencial para a maioria dos organismos. [3]. Os íons de zinco formam complexos com várias proteínas que realizam funções metabólicas vitais. O íon zinco é um componente de pelo menos 300 metaloenzimas que catalisam mais de 50 reações bioquímicas (fisiológicas) diferentes [4, 5] .
No entanto, o zinco é um metal pesado. Pode ser encontrado no grupo IIb da tabela periódica dos elementos, junto com os dois metais tóxicos cádmio e mercúrio. No entanto, o zinco é considerado relativamente não tóxico para os humanos [6]. Este elemento é nocivo apenas em doses excessivas [7].
O LD oral 50 para o zinco é próximo a 3 g / kg de peso corporal, de acordo com o banco de dados TOXNET da US National Library of Medicine. É mais de 10 vezes maior do que o cádmio e 50 vezes maior do que o mercúrio [6]. O excesso da concentração normal desse microelemento em humanos é mais comumente causado pela ingestão de medicamentos e aditivos biologicamente ativos contendo zinco redundante em sua composição. Foi registrado como casos individuais de intoxicação por zinco como resultado da ingestão de alimentos armazenados em recipientes revestidos ou totalmente de zinco. Óxido de zinco, cloreto e sulfato de zinco são amplamente usados na indústria para a produção de vidro; na fabricação de fibras artificiais, tintas de zinco, cerâmicas, fósforos e cimento dentário; na indústria de celulose e papel, para preservação de madeira, e para estanhagem e solda.
Uma alta concentração de ingestão de zinco alterou a resposta imunológica [8]. Os níveis elevados de Zn, Al, Cu e Fe no cérebro podem facilitar o desenvolvimento ou progressão da doença de Alzheimer de acordo com alguns estudos epidemiológicos [9, 10].
Os metais pesados podem afetar a reprodução feminina em diferentes estágios, como o início da vida fetal, desenvolvimento inicial e maturação. Cátions de metais pesados também podem ser a causa de subfertilidade, infertilidade, retardo de crescimento intrauterino, abortos espontâneos, malformações, defeitos de nascença, morte pós-natal, envelhecimento prematuro e distúrbios de aprendizagem e comportamento [11, 12].
A função contrátil uterina está associada à atividade do complexo proteico - actomiosina - no qual a miosina exibe atividade enzimática, ou seja, a capacidade de hidrolisar ATP. A miosina ATPase localizada no domínio catalítico do subfragmento-1 (S1 ou cabeça) transforma a energia química depositada nas ligações macroérgicas do ATP em movimento mecânico. Como resultado, a miosina se move ao longo do filamento de actina, causando a contração muscular. Portanto, a hidrólise do ATP catalisada pela miosina é considerada um dos processos essenciais no mecanismo molecular da função miometrial [13, 14].
O subfragmento-1 da miosina é uma parte N-terminal da cadeia pesada da miosina que consiste em dois domínios:o domínio motor globular N-terminal (catalítico) contendo o local ATP-ase e o local de ligação à actina, e o domínio regulatório, ou braço de alavanca responsável pelo movimento da miosina ao longo dos filamentos de actina. O núcleo do domínio motor da miosina é formado por uma folha β central de sete cadeias que é rodeada por hélices α. Um grande domínio estrutural responsável por seis das sete vertentes da folha β central é geralmente referido como o domínio superior de 50 kDa (U50). Uma grande fenda separa o domínio de 50 kDa superior do domínio de 50 kDa inferior estrutural bem definido (L50) que é formado por resíduos de aminoácidos de 465 a 590. A região de ligação à actina e o local de ligação de nucleotídeo da miosina estão em lados opostos da folha β de sete cadeias com a porção fosfato do nucleotídeo na parte traseira da bolsa de ligação de nucleotídeo. O P-loop, switch 1 e switch 2 estão localizados no domínio superior de 50 kDa próximo ao ápice da grande fenda. Todos os três motivos de ligação de nucleotídeo contatam com a porção fosfato do nucleotídeo na parte traseira da bolsa de ligação de nucleotídeo e agem como sensores de γ-fosfato [15].
Foi descoberto em nossos estudos anteriores que os cátions de metais pesados inibiram a atividade da miosina ATPase do músculo liso uterino [16, 17] que pode afetar negativamente as propriedades contráteis do miométrio.
O impacto adverso do metal pesado na contratilidade uterina requer o desenvolvimento de substâncias farmacológicas que possam eliminar esses efeitos nocivos.
Calixarenes atualmente têm chamado a atenção de pesquisadores como potenciais efetores artificiais para diferentes processos bioquímicos. Estes compostos são oligômeros de fenol macrocíclicos sintéticos que possuem uma estrutura em forma de copo. Suas bordas superior e inferior podem ser funcionalizadas com vários substituintes químicos. Os arenos Calix [4] são formados por quatro fragmentos de arenos funcionalizados e caracterizados por uma conformação macrociclo bastante flexível. Calix [4] arenos apresentou baixa toxicidade da matriz e capacidade de penetração nas células. Portanto, esses compostos são considerados agentes promissores para o desenvolvimento de novos fármacos eficazes [18, 19].
Uma classe promissora de tais substâncias são os tiacalixarenos solúveis em água [18] que possuem os grupos de complexação de metais na plataforma molecular macrocíclica. Os calixarenos devido à sua capacidade de formar os complexos supramoleculares com cátions (bio) metálicos também têm sido utilizados em pesquisas biomédicas como extratores de metais pesados [20,21,22].
Mostramos anteriormente que o tetrahidroxi-tiacalix [4] areno-tetrassulfonato (С-798) eliminou os efeitos inibitórios do Pb 2+ , Cd 2+ e Ni 2+ na hidrólise de ATP catalisada pela miosina S1 do miométrio de suínos [23].
Este estudo teve como objetivo pesquisar o efeito de altas concentrações de cátions zinco e sua ação conjunta com tetrahidroxitiacalix [4] areno-tetrassulfonato (С-798) e tetrahidroxitiacalix [4] areno-tetrafosfonato (C-800) na miosina Atividade S1 ATPase do útero. Este estudo foi necessário para testar a capacidade desses tiacalixarenos de eliminar os efeitos adversos de altas concentrações de zinco na atividade enzimática da miosina uterina.
Thiacalix [4] arenos C-798 e C-800 consistem em um copo formado por quatro fragmentos fenólicos modificados na borda superior com quatro grupos sulfonato aniônico e quatro grupos fosfonato, respectivamente. Ambos os arenos thiacalix [4] têm grupos hidroxila e átomos de enxofre bivalentes densamente localizados na borda inferior, permitindo que eles quelem metais pesados com a formação de complexos de metal estáveis [21] (Fig. 1).
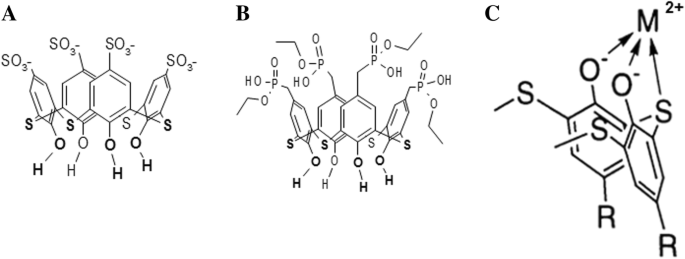
A estrutura química do tetrahidroxi-tiacalix [4] areno-tetrassulfonato (C-798) ( a ), tetrahidroxi-tiacalix [4] areno-tetrafosfonato (C-800) ( b ), e o esquema do complexo quelante do tiacalixareno com cátion metálico na borda inferior (posição invertida) ( c )
Este trabalho é resultado de um projeto conjunto Palladin Institute of Biochemistry e Institute of Organic Chemistry de NAS da Ucrânia focado na interação da miosina ATPase do miométrio com calix [4] arenos que são inibidores ou ativadores (efetores) da miosina ATPase uterina.
Resultados
Dependência da atividade de Miosina S1 ATPase em Zn 2+ Concentração
Verificou-se que o efeito inibitório mais pronunciado na atividade da miosina S1 ATPase do útero foi de 5 mM (43 ± 8%, M ± SD) para cátions Zn. A faixa de concentração de Zn 2+ foi de 0,5-5 mM no meio de incubação (contendo ATP 3 mM, Mg 5 mM 2+ e 0,01 mM Ca 2+ ) Cem por cento é o valor da atividade ATPase sem a adição de cátions Zn (controle) (Fig. 2). Assim, os efeitos adversos dos cátions Zn na hidrólise S1 ATP da miosina foram estudados posteriormente com 5 mM de Zn 2+ .
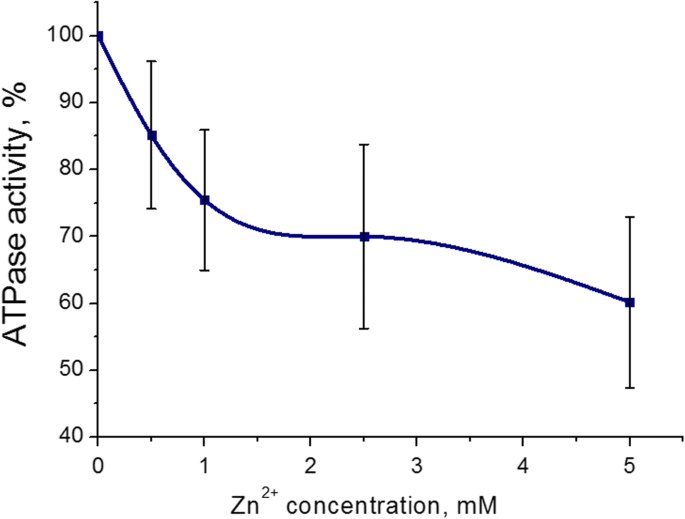
Atividade de miosina S1 ATPase do miométrio na presença de concentrações de 0,5-5,0 mM de cátions Zn (M ± SD, n =6). 100% é o valor da atividade ATPase sem a adição de cátions Zn
Dependência da atividade da miosina S1 ATPase na concentração de ATP na presença 5 mM Zn 2+
O Zn 2+ foi investigado o efeito sobre a afinidade da atividade ATPase S1 da miosina para o seu substrato (ATP). Aumentando a concentração de ATP no meio de incubação de 0,5 a 5 mM em um MgCl fixo 2 concentração (5 mM) tanto no controle quanto na presença de 5 mM de Zn 2+ resultou em um gráfico em forma de cúpula com um valor máximo de atividade de ATPase a 3 mM de ATP. O valor da atividade enzimática neste pico na presença de zinco foi 30% inferior ao do controle (Fig. 3). Os gráficos da dependência da atividade ATPase S1 da miosina na concentração de ATP no controle e a presença de Zn 5 mM 2+ na parte ascendente foram linearizados de acordo com o método Lineweaver-Burk [27]. O cálculo dos parâmetros cinéticos, nomeadamente a constante imaginária de Michaelis ( K m ) e taxa máxima de miosina S1 ATPase para ATP ( V max, ATP ) revelou que V max, ATP da atividade enzimática da miosina na presença de 5 mM de Zn 2+ diminuiu 1,6 vezes (38 ± 7 e 22 ± 6 nmol Pi / min por 1 mg de proteína no controle e a presença de Zn 2+ respectivamente, n =5). O valor de К m para a hidrólise de ATP pela miosina S1 não muda estatisticamente, embora tenda a diminuir (0,49 ± 0,15 mM no controle, 0,38 ± 0,12 mM na presença de Zn 2+ ; M ± SD; n =5).
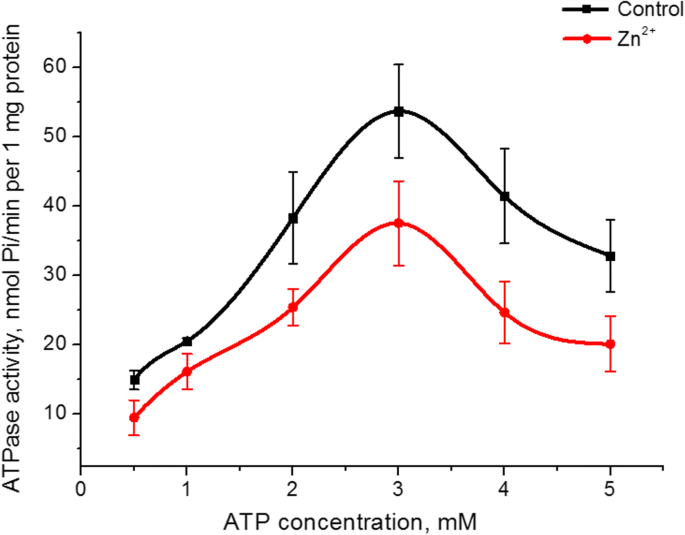
A influência de 0,5-5 mM ATP na atividade da miosina S1 ATPase do útero na presença de 5 mM de Zn 2+ em comparação com um controle (M ± SD, n =5)
Dependência da atividade da Miosina S1 ATPase em Mg 2+ Concentração na presença de 5 mM Zn 2+
O efeito de 5 mM Zn 2+ em Mg 2+ a dependência da concentração na atividade ATPase da miosina uterina foi estudada. Aumentando o Mg 2+ concentração no meio de incubação de 0,5 a 5 mM a uma concentração de ATP fixa (3 mM) na presença de Zn 5 mM 2+ não resulta em uma alteração da atividade da miosina S1 ATPase. Ao mesmo tempo, o Mg 2+ foi detectada dependência da concentração da atividade ATPase no controle (nas condições padrão). O nível mais alto de hidrólise de miosina ATP no controle foi alcançado com 3 mM de Mg 2+ (Fig. 4). Portanto, a atividade da enzima miosina S1 do útero não depende da concentração de Mg 2+ na presença de altas concentrações de Zn (5 mM).
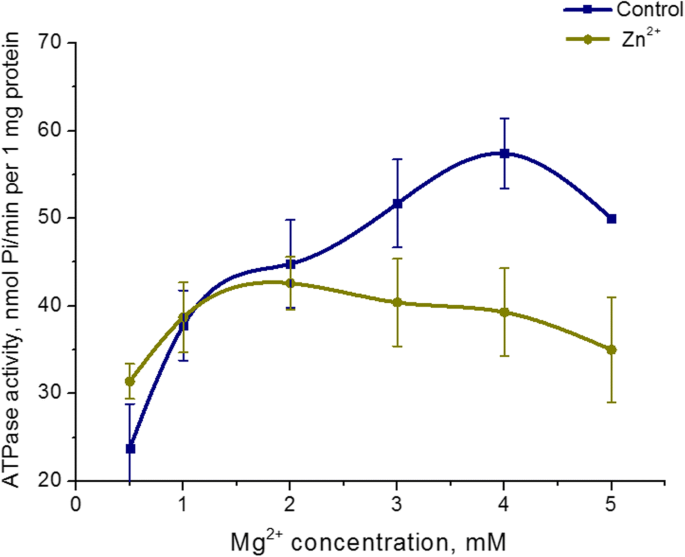
Dependência da atividade da miosina S1 ATPase em Mg 2+ concentração na presença de 5 mM de Zn 2+ em comparação com um controle (M ± SD, n =6)
Zn-Binding Sites em Myosin S1
A simulação de computador mostra que os cátions Zn têm várias regiões de ligação na cabeça da miosina. Um deles está localizado na parte inferior da fenda entre os subdomínios superior e inferior de 50 kDa, próximo ao sítio de ligação do nucleotídeo e diretamente próximo ao P-loop. Zn 2+ é coordenado com os átomos de oxigênio Glu177 (comprimento da ligação 0,23 e 0,39 nm), com o átomo de oxigênio Ser178 (comprimento da ligação 0,31 nm) e Arg236 (comprimento da ligação 0,32 nm).
Outra região de ligação de Zn está localizada na parte inferior do subdomínio superior de 50 kDa (Leu218-Asp463, Glu605-Phe621) e perto do switch 1 (Gly233-Phe246) e P-loop. Zn 2+ pode coordenar com o átomo de oxigênio Glu327 (comprimento de ligação 0,21 nm), com um átomo de oxigênio Glu326 (comprimento de ligação de 0,34 nm) e um átomo de oxigênio Asp323 (comprimento de ligação 0,32 nm). O cátion Zn também pode interagir com a miosina S1 na região que contata o switch 2, interagindo com Glu 465 (0,24 nm), Asp468 (comprimento de ligação 0,31 nm) e Leu653 (comprimento de ligação 0,37 nm). Esta região de ligação está perto do local de ligação da actina e da fenda entre os subdomínios superior e inferior de 50 kDa. A parte inferior desta fenda está localizada na bolsa de ligação de ATP. Estes Zn de ligação 2+ Os domínios S1 da miosina desempenham um papel essencial na ligação e hidrólise do ATP. Essas regiões sofrem transformações conformacionais complexas no processo de transferência de energia do local de hidrólise do ATP para a superfície de ligação da actina.
Thiacalix [4] arenos elimina os efeitos inibitórios do Zn 2+ na atividade da miosina ATPase
Cem micromolares de soluções C-800 ou C-798 em tampão Tris-HCl 50 mM (pH 7,2) foram adicionados ao meio de incubação contendo cátions Zn 5 mM para remover o impacto negativo de Zn 2+ sobre a atividade ATPase da miosina S1 do músculo liso uterino. Como controle, foi utilizado como atividade enzimática sem adição de zinco e / ou tiacalix [4] arenos ao meio de incubação. Foi mostrado (Fig. 5) que o composto C-800 não afeta a atividade ATPase da miosina S1 do miométrio. Embora, o composto C-798 exiba um pequeno efeito inibidor na atividade ATPase S1 da miosina que está provavelmente relacionado à extração de uma certa quantidade de Mg 2+ [21], essencial para a ligação do ATP no centro ativo e sua hidrólise, a partir do meio de incubação. No entanto, 100 μM C-798, bem como C-800, removeram os efeitos inibitórios de 5 mM Zn 2+ no processo de hidrólise de ATP catalisada pela miosina S1.
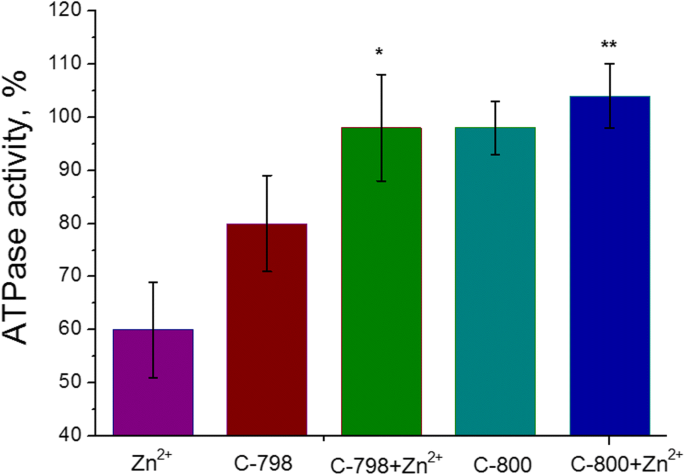
Efeito de 100 μM de C-798 e C-800 na atividade de miosina S1 ATPase na presença de 5 mM de Zn 2+ (M ± SD, n =5-6). 100% é o valor da atividade ATPase sem a adição do cátion Zn. A diferença entre "Zn" e "Zn + C-798", bem como entre os valores de "Zn" e "Zn + C-800", é estatisticamente significativa ( p <0,05) e mostrado como * e **, respectivamente
Prováveis mecanismos de ação restauradora de C-798 e C-800 na atividade de miosina S1 ATPase na presença de Zn 2+
Um dos mecanismos mais prováveis da ação restauradora de C-798 e C-800 na atividade da miosina S1 ATPase em Zn 2+ presença pode ser a capacidade do thiacalix [4] arenos de ligar cátions Zn e, conseqüentemente, excluir esses cátions do meio de incubação. Foi interessante se esses thiacalix [4] arenos podem se ligar a cátions de zinco que já estão ligados à miosina.
A simulação computacional mostrou que os thiacalix [4] arenos C-798 e C-800 com ponte de átomos de enxofre entre anéis aromáticos estão na conformação de “cone” estabilizado por ligações intra-hidrogênio entre os grupos fenólicos. A estrutura de energia minimizada desses calixes [4] arenos foi obtida. A energia total do C-798 após a minimização foi de 64,5 kcal / mol. A presença de grupos ionizados nas bordas calix [4] areno (em particular as inferiores) aumenta significativamente a contribuição das interações eletrostáticas para a energia total da interação hospedeiro-hóspede. Também realizamos a “minimização” do C-798 — Zn 2+ complexo; sua energia total era de 83 kcal / mol.
C-798 foi incorporado na estrutura da miosina S1 cooperando com o cátion Zn que foi previamente ligado à proteína na região do loop P. Neste caso, Zn 2+ interage com os átomos de oxigênio da borda inferior e enxofre de ponte de C-798 (O3, 0,21 nm; S1, 0,30 nm; O2, 0,34 nm). É mostrado que Zn 2+ desvia em certa medida dos resíduos de aminoácidos do P-loop e enfraquece sua interação com o átomo de oxigênio Glu177 (comprimento de ligação de 0,43 nm) (Fig. 6).
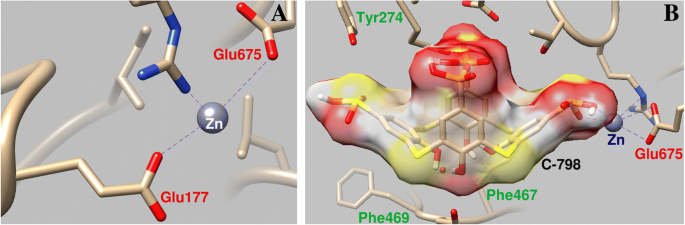
Parâmetros geométricos da interação de Zn 2+ com a região P-loop da miosina S1 ( a ) e a influência de C-798 na cooperação do cátion Zn com esta região ( b )
A fixação do C-798 na “cavidade” da região de ligação do ATP da miosina ocorre com a participação de vários resíduos de aminoácidos. Em particular, a cesta hidrofóbica de tiacalixareno foi fixada pelos resíduos de aminoácidos aromáticos de miosina de Phe467 e Phe469; átomos de oxigênio de tiacalixareno carregados negativamente interagem com resíduos de aminoácidos carregados positivamente de Arg570, Asn572 e His689.
O estudo da influência do C-798 na mudança do Zn 2+ posição durante o encaixe na área perto do local de ligação do ATP da miosina demonstrou que Zn 2+ na presença de tiacalix [4] areno interage com os átomos de oxigênio do terceiro grupo sulfonil (O16–0,26 nm; O15–0,27 nm), quase não interage com os átomos de oxigênio Asp134 e Glu326, e a coordenação com Glu327 é muito mais fraca (Comprimento de acoplamento de 0,43 nm) (Fig. 7). Nesse caso, o tiacalixareno é fixado na “cavidade” da proteína com a participação de vários resíduos de aminoácidos. Em particular, átomos de oxigênio carregados negativamente de grupos tiacalixareno sulfonil interagem com resíduos de aminoácidos carregados positivamente de miosina de Lys188, Lys195 e Gln221.
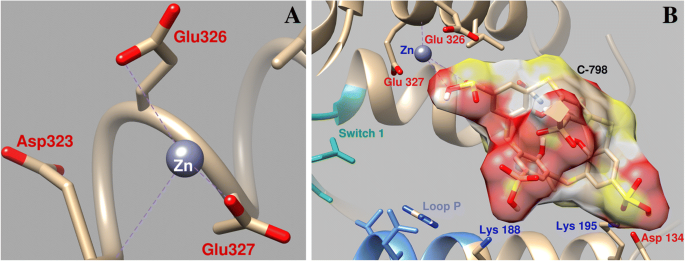
Parâmetros geométricos da interação de Zn 2+ na área perto do interruptor 1 da miosina e da alça P da miosina S1 ( a ) e a influência de C-798 na cooperação de Zn 2+ com esta região ( b )
É mostrado que a esfera de energia condicional do cátion Zn contata e até mesmo se sobrepõe um pouco com a superfície de propagação das interações eletrostáticas devido à presença de átomos de oxigênio e átomos de enxofre em C-798. Isso indica que o cátion sendo estudado está em interação próxima o suficiente com átomos carregados negativamente das coroas C-798 superior e inferior. É provável que o tiacalixareno atraia sobre si mesmo um Zn 2+ , como resultado do qual a interação dos cátions com os resíduos de aminoácidos da enzima é enfraquecida.
Consequentemente, os resultados do acoplamento do C-798 na região da miosina S1, que contém Zn 2+ ligado, indica a possibilidade da interação de grupos funcionais C-798 com o cátion Zn. Neste caso, a ligação do cátion Zn com os resíduos de aminoácidos da miosina S1 foi significativamente enfraquecida, e a distância entre eles aumenta. Como resultado, o efeito adverso do cátion Zn na atividade ATPase da miosina pode ser eliminado.
Também realizamos uma simulação computacional do efeito do calix [4] arene C-800, com docking na área próxima à região de ligação de ATP da miosina S1, para alterar a geometria do cátion Zn. Ao mesmo tempo, Zn 2+ está em contato com Arg236 (comprimento de ligação de 0,37 nm), Glu675 (comprimento de ligação de 0,41 nm) e átomos dos resíduos de calix funcional superior [4] areno (H26, 2,36 nm; H30, 2,96 nm; C30, 0,31 nm; O5, 3,7 nm; O16, 0,4 nm; O7, 0,48 nm; e O11, comprimento de ligação de 0,51 nm). Zn 2+ interage com calix [4] areno C-800, semelhante a C-798. O cátion também é retirado do sítio de ligação anterior nesta região, e está em contato com Asp468 (comprimento de ligação de 0,2 nm) e átomos da coroa de calixareno inferior (C4, 1,96 nm; C14, 2,07 nm; S3, 2,16 nm; O2 , 2,26 nm; C3, 2,97 nm; C20, 3,10 nm; O4, 4,2 nm; C14, 3,0 nm; e O3, 4,1 nm).
O acoplamento do C-798 e do C-800 na região do subfragmento-1 da miosina mostrou que esses tiacalix [4] arenos poderiam interagir com os cátions Zn que se ligavam aos resíduos de aminoácidos da miosina próximos ao sítio ativo da ATPase. Portanto, seu efeito protetor pode ser resultado do enfraquecimento da interação entre esses cátions e a miosina S1.
Discussão
Numerosas anomalias reprodutivas femininas são causadas por distúrbios do músculo liso uterino (miométrio). Os metais pesados têm um efeito adverso na contratilidade do músculo liso uterino. O zinco de metal pesado é um elemento biogênico essencial para a maioria dos organismos; altas doses desse elemento são tóxicas [5]. Várias investigações das concentrações milimolares de Zn 2+ em objetos vivos foram descritos anteriormente [32]. Descobrimos que o Zn 5 mM tem o efeito inibitório mais pronunciado sobre a atividade ATPase S1 da miosina no útero. Assim, os efeitos adversos dos cátions Zn na hidrólise S1 ATP da miosina foram estudados posteriormente com esta concentração de Zn 2+ . O cálculo dos parâmetros cinéticos da miosina S1 ATPase para ATP revelou que V max, ATP da atividade enzimática da miosina na presença de 5 mM de Zn 2+ diminuiu 1,6 vezes. O valor de К m para a hidrólise do ATP não muda estatisticamente, embora tenda a diminuir.
A miosina em condições fisiológicas é um Mg 2+ -dependent ATPase. O cátion magnésio está envolvido na ligação do ATP no sítio ativo da miosina, bem como na hidrólise do ATP. O Mg 2+ é coordenado no sítio ativo da enzima com as cadeias laterais dos resíduos de aminoácidos da miosina Thr-186 e Ser-237, bem como grupos β- e γ-fosfato da molécula de ATP com a formação de β- e γ- complexo bidentado, bem como com moléculas de água ativas, uma das quais realiza um ataque nucleófilo no γ-fosfato ATP [33, 34]. Mg 2+ interage com grupos de fósforo ATP carregados negativamente, polariza-os e, assim, facilita um ataque nucleofílico no γ-fosfato terminal [14].
Verificou-se que a atividade da miosina S1 ATPase não é sensível à presença de Mg 2+ na concentração de 5 mM de Zn 2+ em contraste com o controle quando o zinco estava ausente no meio de incubação [35, 36].
A atividade da miosina ATPase depende da natureza do cátion metálico e se correlaciona bem com seu raio iônico. Os raios de íons Mg 2+ e Zn 2+ nas soluções são muito semelhantes (0,070 e 0,076 nm, respectivamente) [37]. Portanto, a interação de Zn 2+ cátions com Mg 2+ - locais de ligação de miosina são possíveis. Assim, o Mg 2+ - os locais de ligação podem ser ocupados por Zn 2+ cátions em suas altas concentrações. A atividade ATPase da miosina S1 em tais condições pode ser insensível aos cátions de magnésio. A miosina contém dois locais de alta afinidade para Mg 2+ e Mg 2+ ligado a esses locais tem um papel fisiológico importante no processo de transdução de energia durante a contração muscular. Ainda existem vários Mg 2+ - sítios de ligação além do sítio ATPase na molécula de miosina, diferindo na energia de ligação dos íons de magnésio e sua afinidade [35]. Portanto, pode-se assumir que Zn 2+ também pode se ligar a outros locais funcionalmente importantes da miosina S1 que afetam a ligação e a hidrólise do ATP.
A simulação de computador mostra que os cátions Zn têm várias regiões de ligação na cabeça da miosina localizadas perto do local de ligação do ATP, a saber, P-loop e switch 2 de subdomínios superior e inferior de 50 kDa. Esses Zn de ligação 2+ Os domínios S1 da miosina desempenham um papel essencial na ligação e hidrólise do ATP. Essas regiões sofrem transformações conformacionais complexas no processo de transferência de energia do local de hidrólise do ATP para a superfície de ligação da actina.
Os resultados da análise obtidos por Zn 2+ o acoplamento à miosina S1 indica que um papel chave na ligação deste cátion à molécula de miosina desempenha sua interação com grupos carregados negativamente dos resíduos de aminoácidos da enzima, em particular, Glu e Asp.
A influência prejudicial das concentrações tóxicas de Zn 2+ cátions na atividade da miosina S1 ATPase requer a busca por compostos farmacológicos que possam eliminar a ação desse metal. Os objetos de nosso estudo foram tetrahidroxitiacalix [4] areno-tetrassulfonato (C-798) e tetrahidroxitiacalix [4] areno-tetrafosfonato (C-800), que são capazes de quelar metais de transição e pesados com a formação de complexos metálicos estáveis (Fig. 1). A coroa macrocíclica superior de C-798 e C-800 contém quatro grupos sulfonato aniônico ou quatro fosfonatos, respectivamente, que fornecem boa solubilidade em água de tiacalixareno e adesão a moléculas de proteína devido a contatos eletrostáticos com átomos de nitrogênio carregados positivamente de fragmentos de aminoácidos [21 ]
A ancoragem do C-798 e do C-800 na região S1 da miosina mostrou que esses tiacalix [4] arenos poderiam interagir com os cátions Zn que se ligavam aos resíduos de aminoácidos da miosina próximos ao sítio ativo da ATPase. Portanto, seu efeito protetor pode ser resultado do enfraquecimento da interação entre esses cátions e a miosina S1. Especulou-se que os resultados obtidos poderiam ser utilizados em pesquisas futuras com o objetivo de utilizar este tiacalix [4] arenos como compostos farmacológicos no caso de intoxicações por altas concentrações de zinco.
Conclusões
A alta concentração (5 mM) de cátion Zn inibiu a atividade da miosina S1 ATPase do útero. A influência inibida do Zn está relacionada com uma diminuição da velocidade máxima do ATP de hidrólise catalisada pela miosina S1 na presença de Zn 5 mM 2+ . O valor de К m para ATP não muda estatisticamente, embora tenda a diminuir.
Tetrahidroxitiacalix [4] areno-tetrassulfosfonato (C-798) e tetrahidroxitiacalix [4] areno-tetrafosfonato (C-800) restaurou a atividade da miosina S1 ATPase para o nível de controle na presença de 5 mM Zn 2+ .
Os cátions Zn têm várias regiões de ligação na miosina S1 localizadas perto do sítio ativo da ATPase. O acoplamento de C-798 e C-800 na região da miosina S1, que contém Zn 2+ ligado, indica a possibilidade da interação desses grupos funcionais tiacalix [4] areno com cátions Zn ligados. A ligação do cátion Zn com os resíduos de aminoácidos da miosina S1 foi significativamente enfraquecida e a distância entre eles aumenta. Como resultado, o efeito adverso do cátion Zn na atividade ATPase da miosina pode ser eliminado.
Supõe-se que os resultados obtidos poderão ser utilizados em pesquisas futuras com o objetivo de utilizar este tiacalix [4] arenos como compostos farmacológicos no caso de envenenamento por altas concentrações de zinco.
Métodos
Reagentes
Os seguintes reagentes foram usados:albumina sérica, EGTA, EDTA, ATP, ácido ascórbico, Tris, tricina, ditiotreitol, acrilamida, (Sigma, EUA), glicina (Merck, Alemanha), N, N′-metilenobisacrilamida (Acros Organics, Bélgica ) N, N, N′, N′-tetrametilenodiamina (Reanal, Hungria), e reagentes de produção doméstica (grau R). As soluções foram preparadas em água purificada no sistema Crystal Bio (Adrona, Letônia). A condutância da água era inferior a 0,1 μS. A concentração dos cátions metálicos bivalentes em solução foi determinada pelo método de Mohr.
Isolamento de Actomiosina e Subfragmento-1 de Miosina
A actomiosina foi isolada do músculo liso uterino de suínos pelo método de Barany modificado, conforme descrito em [17]. A miosina S1 foi obtida da actomiosina suína pelo método de Suzuki modificado [24]. A pureza das amostras foi controlada por eletroforese PAAG-SDS [25].
Ensaio de atividade de ATPase
ATPase activity of myosin S1 was determined in a 96-well plate at 37 °C in an incubation medium (total volume 0.1 ml) of the following composition (mM):Tris-HCl buffer (pH 7.2), 20; KCl, 100; CaCl2 , 0.01; MgCl2 , 5; and ATP, 3 (standard conditions). Protein (myosin S1) concentration was 20 μg/ml. Incubation time was 5 min. Samples containing all components of the incubation medium without myosin S1 were taken as control of non-enzyme hydrolysis of ATP. The amount of inorganic phosphate released during ATP hydrolysis reaction was determined by the Chen method [26] by the measurement of optical absorbance of the solution at 820 nm using a microplate reader μQuwant (Biotek @ Instruments, Inc., USA) and specified as Pi nmol/min per 1 mg of protein.
The Zn 2+ and thiacalix[4]arene effects on the ATPase activity of myosin S1 were studied using standard incubation medium with solutions of ZnCl2 and thiacalix[4]arenes at the corresponding concentrations. The value of ATPase activity in the absence of ZnCl2 and/or calix[4]arenes in the incubation medium was taken as 100% (control).
Kinetic and Statistical Analysis
The values of the imaginary constant of Michaelis (K m ) and maximal rate of myosin S1 ATPase for ATP (V max, ATP ) were calculated using the graph of the dependence of ATPase activity on the ATP concentration according to Lineweaver–Burk method [27]. Statistical processing of the obtained data was performed using standard methods of variation statistics. Experimental data were analyzed by using the standard software “MS Office” and “Statistica 4.5.” The statistical comparisons were performed using two-way analysis of variances (ANOVA).
Thiacalix[4]Arene Synthesis and Characterization
Tetrahydroxy-thiacalix[4]arene-tetrasulphonate and tetrahydroxy-thiacalix[4]arene-tetraphosphonate were synthesized and characterized using NMR techniques and IR spectroscopy in the Phosphoranes Chemistry Department of the Institute of Organic Chemistry, NAS of Ukraine. Infrared and NMR spectroscopy confirmed the structure of these synthesized thiacalix[4]arenes. This thiacalix[4]arenes were dissolved in water.
Computer Modeling
Computer modeling of the interaction between ligands (thiacalix[4]arenes, Zn 2+ , model bindings) and receptor (myosin S1) was performed using AutoDock software, version 4.2 [28]. We used the three-dimensional enzyme structure with the 1b7t identifier in RSCB PDB in our research [29]. Computer modeling of the thiacalix[4]arene structural peculiarities was carried out using HyperChem 7.01. Molecular dynamics calculations were performed by the MM2 method with the semi-empirical methods (CNDO).
Program AutoDockTools was used for preliminary “processing” of interacting molecules. One hundred runs of Lamarkian genetic algorithms (population size, 100; the maximal number of energy evaluations, 10 6 ) were conducted. To analyze and visualize the docking results, we used the programs Chimera [30] and Yassara [31]. Calculation of the minimal total binding energy was implemented considering Van der Waals forces, electrostatic and hydrophobic interactions, and hydrogen bonds. The optimal ligand positions in the complex “receptor-ligand” were determined according to the energy values obtained by docking software calculator for binding energy in complex “receptor-ligand.” Thus, we selected a series of complexes with the lowest total energy and then calculated the optimal geometry of the complexes and determined the most energetically preferred arrangement of the ligands in the space of myosin subfragment-1 binding domain.
Abreviações
- C-798:
-
Tetrahydroxythiacalix[4]arene-tetrasulfosphonate
- C-800:
-
Tetrahydroxythiacalix[4]arene-tetraphosphonate
- CNDO:
-
Complete Neglect of Differential Overlap (methods)
- K m :
-
Michaelis constant, the substrate concentration at which the reaction rate of the enzyme is half of the maximal velocity
- L50:
-
Lower 50-kDa domain of myosin
- LD50 :
-
Lethal dose is the amount of an ingested substance that kills 50% of a test sample
- MM2:
-
A class of force fields
- Myosin S1:
-
Myosin subfragment-1
- NASU:
-
National Academy of Science of Ukraine
- PDB:
-
Protein Data Bank
- P-loop:
-
Phosphate-binding loop of myosin
- RCSB:
-
Research Collaboratory for Structural Bioinformatics
- U50:
-
The upper 50-kDa domain of myosin
- V máximo :
-
Maximal velocity of the enzyme
- V max, ATP :
-
V máximo for ATP
Redistribuição do campo eletromagnético em nanopartículas de metal sobre grafeno
Anisotropia Magnética Perpendicular e Mudança Magnética Induzida por Hidrogenação de Multicamadas Ta / Pd / CoFeMnSi / MgO / Pd
Nanomateriais
- Aço Zincado vs. Aço Galvanizado:Qual é a Diferença?
- Os efeitos do COVID-19 na manufatura
- A administração de nanopartículas de artesunato aumenta a eficiência antitumoral ao ativar a apoptose celular mediada por mitocôndrias
- Os efeitos da relação Li / Nb na preparação e desempenho fotocatalítico de compostos Li-Nb-O
- Os efeitos positivos da automação agrícola
- Quedas de energia:5 maneiras de evitar os efeitos negativos
- Os efeitos do impulso radial em bombas centrífugas
- Os efeitos de queima de plasma e oxi/combustível
- Os efeitos dos elementos de liga comuns
- Quais são os efeitos da atual escassez de níquel?