


O papel da apoptose na citotoxicidade induzida por nanopartículas de óxido de zinco frescas e envelhecidas
Resumo
Nanopartículas de óxido de zinco (ZnO NPs) são usadas em uma ampla gama de aplicações, incluindo a indústria, produtos comerciais e área médica. Numerosos estudos mecanísticos para a toxicidade de NPs de ZnO foram realizados em NPs puros (frescos). No entanto, a citotoxicidade induzida pelos NPs de ZnO transformados (envelhecidos) e os mecanismos subjacentes permanecem obscuros. Aqui, observamos a transformação físico-química de NPs de ZnO sofrida ao longo do tempo, seguida pela avaliação da citotoxicidade de NPs frescos e envelhecidos. Descobrimos que NPs de ZnO frescos induziam maior nível de apoptose do que seus equivalentes envelhecidos. Consequentemente, os dados de sequenciamento de RNA de híbrido humano-hamster tratado com ZnO NP envelhecido ( A L ) as células mostraram que as vias de sinalização p53, PI3k – Akt, FoXO, Glutationa, ErbB, HIF-1, oxitocina e Jak-STAT foram enriquecidas, mas nenhuma via de apoptose. Os resultados quantitativos da PCR revelaram o nível de mRNA significativamente mais alto de IL1B e CD69 em grupos tratados com NP fresco em comparação com os grupos tratados com ZnO NP- e cloreto de zinco. Os resultados acima indicaram que a menor citotoxicidade de NPs de ZnO envelhecidos é parcialmente atribuída à sua potência reduzida na indução de apoptose. A regulação transcricional de múltiplas vias de sinal ativadas por NPs idosos pode ajudar a construir a homeostase celular. Tomados em conjunto, nossos resultados destacam a influência do processo de envelhecimento (transformação ambiental) de NPs de ZnO em suas toxicidades e consequências biológicas.
Introdução
Com o rápido desenvolvimento da nanotecnologia nas últimas décadas, as nanopartículas (NPs) têm sido aplicadas em vários campos, incluindo a indústria, a vida diária humana e a nanomedicina [1, 2]. O Inventário de Produtos de Consumo de Nanotecnologia (CPI) mostra um aumento de 30 vezes entre 2005 e 2015 no número de nanoprodutos, incluindo 762 produtos de saúde (fitness), 72 alimentos (bebidas) e 23 produtos para bebês [2]. A crescente aplicação de NPs em produtos de consumo e em vários campos aumentou a possibilidade de NPs entrarem no meio ambiente, o que levanta questões de segurança no que diz respeito aos seus potenciais impactos adversos. NPs de óxido de zinco (ZnO) estão entre os NPs mais comumente usados, e sua produção anual global atingiu quase 3400 toneladas [3, 4]. Algumas substâncias antes consideradas biologicamente inertes podem se tornar tóxicas em seu estado nanoparticulado. Um número crescente de estudos elucidou que os NPs de ZnO podem representar riscos significativos para células de mamíferos e animais, induzindo toxicidade significativa [5,6,7].
Várias estratégias, incluindo revestimento, funcionalização de superfície e modificação do estado de oxidação têm sido usadas para atenuar a toxicidade potencial dos NPs, modificando as propriedades físicas e químicas deles (como a dissolução, aglomeração e perturbação das membranas celulares) [8,9,10, 11]. Embora essas modificações de NPs enfraqueçam seus efeitos tóxicos em certo grau, os usos de NPs nem sempre são seguros, especialmente sob certas condições de exposição e ambientes [12,13,14]. Na verdade, muitos tipos de NPs não são estáveis e tendem a sofrer “envelhecimento” ou “transformação ambiental” após serem intencionalmente ou não liberados no ambiente natural [14,15,16,17]. Nos últimos anos, muitos trabalhos foram realizados para explorar o processo de transformação ambiental das NPs; entretanto, a pesquisa sobre os efeitos tóxicos de NPs “transformados (envelhecidos)” ainda é muito limitada, quanto mais seus mecanismos tóxicos.
Como o representante típico de NPs não persistentes, os NPs de ZnO têm reatividade muito alta e são propensos a se transformar em propriedades físicas e químicas e estado de ocorrência após serem liberados no meio ambiente ou ingeridos por animais, o que poderia afetar significativamente seus efeitos toxicológicos [17 , 18]. Por exemplo, estudos mostraram que o processo de sulfetação de NPs de ZnO alterou a carga, a hidrofobicidade e o estado de agregação, resultando na adsorção de NPs em estado de sulfeto na saliva humana, suor e fluido de lavagem broncoalveolar. Além disso, a proteína adsorvida por ZnO NPs forma uma coroa protéica especial que geralmente afeta seu efeito biológico [19]. Os fosfatos em soluções fisiológicas podem converter ZnO NPs em ZnHPO metaestável 4 e Zn 3 (PO 4 ) 2 dentro de cerca de 5-10 h [20]. O processo de transformação completa de ZnO NPs (≤ 3 μg / mL) no sistema de exposição in vitro para linfócitos T humanos (37 ° C, meio de cultura celular RPMI1640 contendo 10% de FBS por 24 h) foi investigado usando radiação síncrotron X- espectroscopia de absorção de raio próximo à borda (XANES) [21]. Os estudos acima sugerem a subestimação dos riscos ambientais e de saúde dos NPs de ZnO, avaliando apenas os efeitos biológicos dos NPs de ZnO puros (frescos). Diante desse problema, há uma necessidade urgente de compreender de forma abrangente os processos de envelhecimento e transformação ambiental das NPs [22].
Nosso estudo anterior revelou que os NPs de ZnO envelhecidos por 40-120 dias em água ultrapura sofrem transformação físico-química e se transformam em Zn 5 (CO 3 ) 2 (OH) 6 , Zn (OH) 2 e Zn 2+ [23]. Curiosamente, os NPs de ZnO envelhecidos exibiram menor citotoxicidade do que as contrapartes frescas [23], mas os mecanismos de toxicidade desse tipo de variação não são claros. No presente estudo, pretendemos explorar as razões subjacentes de diferentes citotoxicidades entre NPs de ZnO frescos e envelhecidos. NPs de ZnO com dois tamanhos de partícula diferentes (20 nm e 90–200 nm) foram aplicados sistematicamente. Os ensaios de citotoxicidade demonstraram que os NPs de ZnO envelhecidos induziram anormalidades morfológicas menos pronunciadas e viabilidades celulares relativamente mais altas do que suas contrapartes frescas. Os dados de sequenciamento de RNA revelaram que os genes apoptóticos foram enriquecidos em células tratadas com ZnO NP frescas, ao passo que esses genes foram muito menos afetados por tratamentos com ZnO NP envelhecidos. Além disso, as células expostas a NPs de ZnO envelhecidos mostraram nível reduzido de proteína Caspase-3 clivada, indicando ainda a maior potência de NPs de ZnO frescos em desencadear apoptose em células cultivadas. Combinado com nossos achados anteriores, este estudo sugeriu que a diminuição da citotoxicidade de NPs de ZnO envelhecidos é atribuída à sua capacidade atenuada em desencadear a apoptose celular.
Materiais e métodos
Nanopartículas e Reagentes
Os nanopós de ZnO comercialmente disponíveis (ZnO NPs), com tamanho médio relatado pelo fabricante de 20 nm (99,5% de pureza, quase esférico) e 90-200 nm (99,9% de pureza, morfologia irregular), foram adquiridos de Nanostructured &Amorphous Materials (Houston, TX ) Exceto quando indicado de outra forma, todos os reagentes e produtos químicos usados neste estudo foram adquiridos da Sigma-Aldrich (Shanghai, China).
Dispersão, envelhecimento e caracterização de nanopartículas
As suspensões de estoque de ZnO NPs (1 mg / mL) foram preparadas pela suspensão de nanopós secos em Milli- Q H 2 O (Millipore, 18 MΩ cm) e esterilizado em autoclave (120 ° C, 30 min) e em seguida armazenado a 25 ° C por período de envelhecimento natural variando de 0 a 60 dias. Os NPs de ZnO naturalmente transformados de 0 e 60 dias foram designados como NPs frescos e envelhecidos, respectivamente. Para garantir a dispersão adequada, as suspensões frescas e envelhecidas foram sonicadas (100 W) por 30 min em um banho ultrassônico antes da caracterização ou incubação com as células. A morfologia, o tamanho de partícula e a agregação de NPs de ZnO frescos e envelhecidos foram caracterizados por meio de microscopia eletrônica de transmissão (TEM, JEOL JEM-2010, Tóquio, Japão). A estrutura cristalina de ZnO NPs frescos e envelhecidos foi determinada usando difração de raios-X de potência (XRD, PANalytical B. V., Shanghai, China) por comparação com padrões autênticos. Os detalhes do processo natural de envelhecimento e caracterização em NPs de ZnO foram descritos anteriormente [23].
Cultura de células e tratamento com NPs de ZnO
A L linha celular, um tipo de células híbridas humano-hamster formadas pela fusão do gly2A mutante de ovário de hamster chinês (CHO) e fibroblastos humanos foi usado neste estudo. Estas células híbridas continham um conjunto padrão de cromossomos CHO-K1 e uma única cópia do cromossomo humano 11 e foram cultivadas em meio F12 de Ham (Hyclone, Grand Island, NY) suplementado com soro fetal bovino (8%, Hyclone, Grand Island, NY) ), gentamicina (25 g / mL) e glicina (2 × 10 –4 M) a 37 ° C em um umidificado 5% CO 2 incubadora [24]. As suspensões de estoque de NPs de ZnO frescas e envelhecidas foram dispersas por 30 min de ultrassonicação (100 W) para evitar aglomeração, posteriormente diluídas para concentrações adequadas com meio de cultura de células para a exposição das células. As células mantidas em meio de cultura celular sem NPs foram servidas como controle em cada experimento.
Ensaio para detectar a citotoxicidade
A L as células em uma fase logarítmica de crescimento foram cultivadas em lâminas de vidro em placas de Petri de 35 mm (6 × 10 4 células / placa) por 24 h antes da estimulação, seguido por tratamento com 2 mL de meio de cultura contendo 1, 5, 10, 12, 15 e 20 µg / mL de ZnO NPs frescos ou envelhecidos 72 h. Após o término do tempo de tratamento, as imagens da morfologia celular foram obtidas em um microscópio Leica DM4B (Leica, Alemanha). ZnCl 2 foi incluído como referência de íons zinco para comparar a citotoxicidade com ZnO NPs.
O kit de contagem de células (CCK-8) (APExBIO, Shanghai, China) foi utilizado para detectar a viabilidade celular. Em detalhes, A L as células foram semeadas em placas de 96 poços (4 × 10 3 células / poço) com meio de cultura de células por 24 horas e tratadas com meio contendo várias concentrações de ZnCl 2 , NPs de ZnO envelhecidos e frescos por 24, 48 e 72 h, respectivamente. Para a solução de trabalho, o volume de NPs adicionados da suspensão de estoque foi inferior a 5% do volume total do meio de cultura em cada poço. Após a conclusão do tempo de tratamento, o meio de cultura foi aspirado e as células foram incubadas com 100 µL de solução de trabalho CCK-8 por 2 h a 37 ° C seguindo as instruções do fabricante. Em seguida, a absorbância foi registrada a 450 nm usando um leitor de fluorescência Spectra Max M2 (Molecular Devices, Wokingham, Berks, UK). A viabilidade celular foi calculada como uma porcentagem da absorbância nos poços, com cada concentração de NPs normalizada para a absorbância das células controle (100%).
Extração de RNA, transcrição reversa e PCR quantitativo
A L células em uma fase logarítmica de crescimento foram semeadas em placas de Petri de 35 mm de diâmetro (6 × 10 4 células / placa) com meio de cultura de células por 24 h. Em seguida, o meio foi substituído por 2 mL de meio de cultura contendo 12 µg / mL de ZnCl 2 , NPs de ZnO envelhecidos e frescos por 72 h. Após a conclusão do tempo de tratamento, o meio de cultura foi aspirado e as células foram lavadas 3 vezes com PBS. Posteriormente, 1 mL do reagente Trizol (Invitrogen, Carlsbad, CA, EUA) foi adicionado a cada placa para extrair o RNA total de acordo com as instruções do fabricante. A concentração e a pureza do RNA total obtido após a extração foram quantificadas por meio do Espectrofotômetro Q5000UV-Vis (Quawell, EUA). Após a quantificação, a transcrição reversa foi realizada usando o kit TransGene RT-PCR (TransGene Biotech, Pequim, China) para obter cDNA do modelo de RNA de acordo com os protocolos do fabricante. As amostras de cDNA resultantes foram quantificadas usando o espectrofotômetro Q5000 UV-Vis e, em seguida, analisadas usando SYBR-Green como corante de fluorescência (TransGene Biotech, Pequim, China) no sistema Roche RT-PCR (Applied Biosystems) [25].
O gene de manutenção que codifica Gliceraldeído-3-fosfato desidrogenase ( Gapdh ) foi usado como controle interno para avaliar Il-1α , Il-1β , Caspase 3 , CD69 , junho e MT1 Expressão de mRNA. Os resultados foram expressos como a razão de expressão relativa entre o gene alvo e Gapdh . As sequências de primer utilizadas neste estudo são fornecidas na Tabela 1.
Análise de dados de sequenciamento de RNA
O total de amostras de RNA de A L células do grupo de controle, grupo tratado com ZnO NP envelhecido e ZnCl 2 o grupo tratado foi sequenciado pela BangFei Bioscience (Pequim, China). Resumidamente, o RNA total de A L as células foram extraídas seguindo os protocolos TRIZOL, até a precipitação isoproponal. Em seguida, as amostras de RNA foram ressuspensas no tampão de extração antes do sequenciamento. Os dados de sequenciação de RNA de contagem bruta foram analisados usando R package Deseq2 [Eric1]. O diagrama de Venn foi gerado pelo pacote R VennDiagram [Eric1.2]. Os genes significativamente alterados foram usados para análises adicionais de enriquecimento da via. Os experimentos foram realizados em três repetições independentes. Os genes de rRNA, os genes mitocondriais e os genes detectados com menos de 40 pb foram excluídos da análise.
Os dados de sequenciação de RNA, série de referência GSE97852, GSE60159 e GSE39444, foram obtidos de Gene Expression Omnibus [Eric 2, 3, 4]. O gráfico Gene Set Enrichment Analysis foi gerado por R (versão 3.6.2) usando o pacote fgsea [Eric 5]. Os genes de apoptose com mudança significativa de 1,5 vezes & p valor <0,05 foram usados para análises posteriores. O mapa de calor com árvore de genes foi gerado pelo pacote R “ComplexHeatmap” [Eric 6]. A ligação média foi usada como método de agrupamento e euclidiana como método de medição de distância. A análise de enriquecimento da via foi precedida usando STRING2.0 [Eric 7].
Western Blotting
A L células em uma fase logarítmica de crescimento foram semeadas em placas de Petri de 60 mm de diâmetro (1,5 × 10 5 células / placa) com meio de cultura de células por 24 h. Em seguida, o meio foi substituído por 4 mL de meio de cultura contendo 12 µg / mL de NPs de ZnO frescos ou envelhecidos por 24 h. No final do período de exposição, o meio de cultura foi aspirado e, em seguida, as células foram lavadas 3 vezes com PBS e lisadas em gelo com tampão de lise RIPA (Beyotime, China) para coletar as proteínas celulares. Quantidades iguais de proteínas celulares foram separadas em géis de SDS-PAGE a 12% e depois transferidas para uma membrana de fluoreto de polivinilideno (PVDF) (Roche, Swiss). Resumidamente, após 2 h de bloqueio com 5% de leite desnatado em TBST a 25 ° C, as membranas foram subsequentemente incubadas com o anticorpo primário em diluições apropriadas (de acordo com os protocolos do fabricante) a 4 ° C durante a noite, seguido por incubação com HRP-conjugado secundário anticorpos (1:5000, Promega, Madison, EUA) durante 2 h a 25 ° C. Finalmente, a imunomarcação foi detectada usando uma solução de quimioluminescência aprimorada (ECL) (BOSTER, China). Os anticorpos primários de anti-pro / Caspase-3 clivada e anti-Actina foram adquiridos na Cell Signaling Technology e na ImmunoWay, respectivamente.
Estatísticas
A análise estatística foi compilada a partir das médias dos resultados obtidos em pelo menos três experimentos independentes. Todos os dados foram apresentados como média ± desvio padrão (DP) e comparados estatisticamente por meio da análise de variância de um fator (ANOVA). Em todas as parcelas p valores <0,05 foram apresentados como * e considerados estatisticamente significativos.
Resultados
Caracterização de ZnO NPs
Para determinar as diferenças nas características físico-químicas detalhadas entre ZnO NPs frescos e envelhecidos, primeiro observamos a morfologia dos NPs usando TEM (Fig. 1A). Nossos resultados indicaram que os NPs de ZnO frescos de 20 nm eram cristais quase esféricos e os NPs de ZnO frescos de 90-200 nm eram cristais cúbicos / semelhantes a bastonetes. O tamanho de partícula única era consistente com o tamanho fornecido pelo fabricante. Obviamente, ambos os NPs de ZnO de 20 nm e 90-200 nm estavam inclinados a se agregar em água ultrapura. Além disso, independentemente da forma e do tamanho dos NPs originais, a microestrutura dos NPs de ZnO de 20 nm e 90-200 nm mudou drasticamente de uma estrutura cristalina clara para um estado amorfo ou semelhante a uma folha / agulha após envelhecer por 60 dias. Além disso, a natureza cristalina e a pureza de fase de NPs frescas e envelhecidas foram determinadas usando difração de raios-X (XRD) com radiação Cu Kα ( λ =0,15418 nm) abordagem a 25 ° C, como mostrado na Fig. 1B. O padrão de XRD de NPs de ZnO frescos indicou que as amostras eram compostas por uma estrutura wurtzita cristalina e nenhum pico de impureza característico foi identificado, sugerindo uma alta qualidade de NPs frescos. Para NPs envelhecidos, o padrão de XRD exibiu a neoformação de Zn 5 (CO 3 ) 2 (OH) 6 (número do cartão 00-011-0287) e Zn (OH) 2 (cartão número 00-003-0888) fases sólidas, indicando a transformação química de NPs de ZnO (20 e 90–200 nm) durante o processo de envelhecimento.
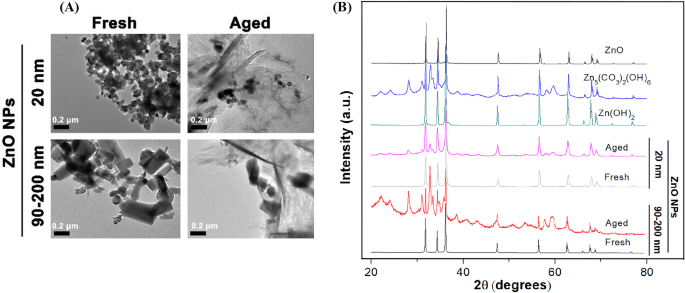
Características físico-químicas de NPs de ZnO frescos e envelhecidos. A Micrografias representativas de NPs frescos e envelhecidos (100 μg / mL, 20 e 90–200 nm) em Milli- Q água usando TEM de baixa resolução, B Padrões de XRD de NPs recentes, NPs envelhecidos, ZnO, Zn (OH) 2 e Zn 5 (CO 3 ) 2 (OH) 6 referências em forma seca
Observação morfológica de A L Células expostas a NPs de ZnO recentes e envelhecidos
O tratamento dos NPs resulta em uma mudança perceptível na forma celular, ou morfologia, in vitro [26]. Portanto, A L as células expostas a NPs de ZnO frescas ou envelhecidas a 10 e 15 µg / mL por 72 h foram examinadas em um microscópio estereoscópico. Conforme mostrado na Fig. 2, a morfologia celular no grupo de controle permaneceu normal. As células aderiram bem, com a maioria aderindo em 2 h. A maioria das células era fusiforme ou poligonal, com algumas células recém-se dividindo apresentando citoplasma mais transparente e melhor dispersão durante o processo de adesão. O tratamento com NPs de ZnO frescos de 12 μg / mL (20 nm e 90–200 nm) por 72 h mudou significativamente a morfologia celular. Embora a maioria das células tenha aderido em 3–5 h, elas não se espalharam bem e algumas células tornaram-se arredondadas e perderam a forma poligonal. Quando a concentração de NPs de ZnO foi aumentada para 15 μg / mL, as células tratadas atrofiaram e não puderam aderir, sugerindo sua viabilidade celular significativamente menor do que a das células tratadas com 10 μg / mL. Esses resultados indicaram que o LC100 para NPs de ZnO frescos é provavelmente inferior a 15 μg / mL por um tratamento de 72 h. Em contraste, a morfologia celular em grupos tratados com NP com idade de 20 nm e 90-200 nm (15 μg / mL) não foi significativamente afetada, e a maioria das células sobreviventes puderam aderir e se espalhar, com menos da metade das células mortas observadas, revelando que NPs de ZnO envelhecidos são muito menos citotóxicos do que NPs de ZnO frescos.
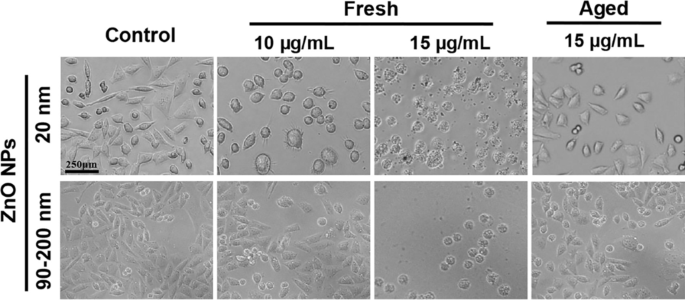
Mudanças morfológicas em A L células após exposição a NPs de ZnO frescas ou envelhecidas por 72 h em meio F12 de Ham suplementar, e células não expostas foram usadas como grupos de controle. A L a morfologia celular foi observada com um microscópio óptico com ampliação de 10 ×
NPs de ZnO envelhecidos induzem citotoxicidade mais baixa do que NPs recentes
Para investigar mais a diferença na citotoxicidade entre ZnO NPs frescos e envelhecidos, examinamos a viabilidade celular usando kits CCK-8. Conforme mostrado na Fig. 3, incubação A L células com doses gradientes de NPs de ZnO frescas e envelhecidas (variando de 0 a 20 μg / mL, 20 nm e 90–200 nm) por 24 h, 48 h ou 72 h mostraram uma diminuição dependente da dose na viabilidade celular. Nenhuma citotoxicidade óbvia foi observada ao tratar células com NPs de ZnO ≤ 10 μg / mL. Quando a dosagem de NPs de ZnO frescos e envelhecidos aumentou para 12 e 15 μg / mL, a viabilidade celular mostrou uma tendência de diminuição dependente do tempo. Obviamente, a viabilidade celular em grupos idosos tratados com NP foi significativamente maior do que grupos tratados com NP fresco. Além disso, ZnCl 2 -tratamento também comprometeu a viabilidade celular de uma maneira dependente da dose e do tempo, enquanto a citotoxicidade do ZnCl 2 era muito menor do que os NPs de ZnO frescos e envelhecidos.
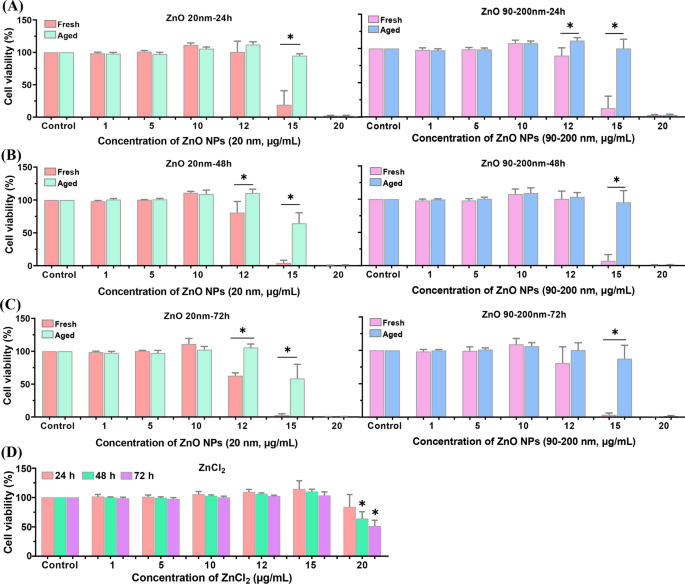
Viabilidade celular induzida por NPs de ZnO frescos e envelhecidos em A L células. A L as células foram incubadas com várias concentrações de NPs de ZnO frescos e envelhecidos (20 e 90–200 nm) por 24 h ( A ), 48 h ( B ) e 72 h ( C ) D A L as células foram expostas a várias concentrações de ZnCl 2 para momentos diferentes. Os dados foram baseados em ≥ 3 experimentos independentes e expressos como média ± DP, * p <0,05
Vias de apoptose ativadas por tratamento com NPs de ZnO frescos e reguladas para cima na expressão de genes apoptóticos
Para desvendar os mecanismos subjacentes que levam à menor citotoxicidade de NPs envelhecidos, analisamos os dados de sequenciamento de RNA de NPs de ZnO novos e envelhecidos. Como mostrado na Fig. 4A, B, após o tratamento com NPs de ZnO frescos, a via de apoptose foi ativada em células Jurkat ( p =0,017) e células HMDM ( p =0,041). Os genes de apoptose: ANXA1 , CYLD , TNFSF10 , IER3 , CDKN1A , JUN , SAT1 , PMAIP1 , CD38 e ISG20 foram significativamente enriquecidos em células Jurkat tratadas com ZnO NP frescas. Os genes de apoptose: CD38 , TNFRSF12A , CCNA1 , BMP2 , PPP2R5B , EREG , IFNGR1 , CD44 , CD14 , GNA15 , GCH1 , TIMP1 , BTG2 , IL1B , IL1A , BTG3 , BCL2L11 , SC5D e SPTAN1 foram significativamente enriquecidos em células HMDM frescas tratadas com ZnO NP (Fig. 4C, D). Como as células de Jurket (linfócitos T do sangue periférico) e as células HMDM (macrófagos derivados de monócitos humanos) são tipos diferentes de células, a maneira como desencadeiam a apoptose pode ser diferente. Em suma, esses resultados mostraram que a exposição a NPs de ZnO frescas pode ativar diferentes vias de apoptose em vários tipos de células.
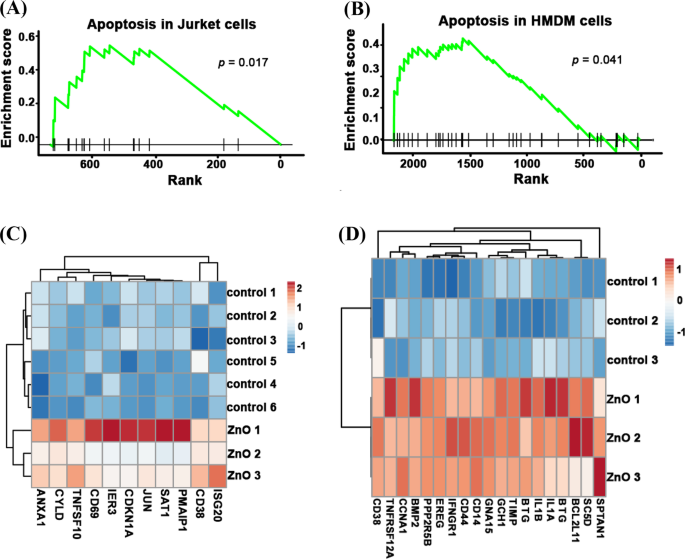
A via de apoptose foi enriquecida em dados de RNA-seq de células Jurket e HMDM tratadas com ZnO NP frescas. A pontuação de enriquecimento de genes expressos de forma significativa da via de apoptose de células Jurket tratadas com ZnO NP frescas ( A ) e células HMDM ( B ) O mapa de calor da expressão gênica apoptótica de células Jurket frescas tratadas com ZnO NP ( C ) e células HMDM ( D ) e seus grupos de controle
NPs de ZnO envelhecidos não regulavam positivamente a expressão de genes apoptóticos como NPs de ZnO recentes
Nossos dados de sequenciamento de RNA de A tratados com ZnO NP envelhecidos L as células mostraram que as vias de sinalização p53, PI3k – Akt, FoXO, Glutationa, ErbB, HIF-1, oxitocina e Jak-STAT foram enriquecidas (Fig. 5A). Os genes de apoptose enriquecidos em células Jurket e HMDM não foram significativamente afetados nas células tratadas com ZnO NP envelhecidas (Fig. 5B). Para confirmar ainda mais os resultados, testamos a expressão de genes relacionados por PCR em tempo real. Descobrimos que alguns dos genes de apoptose: BMP2 , PMAIP1 , IL1α , CD69 , CCNA1 , CD38 e IL1β eram indetectáveis em idosos tratados com NPs de ZnO A L células (dados não mostrados), provavelmente porque a maioria desses genes é expressa em células do sistema imunológico. Os outros genes de apoptose regulados positivamente ( IL1α , IL1β e CD59 ) observados em grupos tratados com ZnO NP fresco não foram significativamente alterados nos níveis de expressão pelo tratamento com ZnO NPs envelhecidos. Enquanto o MT1 que servem como um controle positivo foi significativamente aumentado em um nível de expressão, a expressão de Caspase 3 não foi alterado significativamente (Fig. 5C). Esses dados sugeriram que os NPs de ZnO envelhecidos, ao contrário de suas contrapartes recentes, são menos potentes na ativação de genes da via de apoptose em A L células.
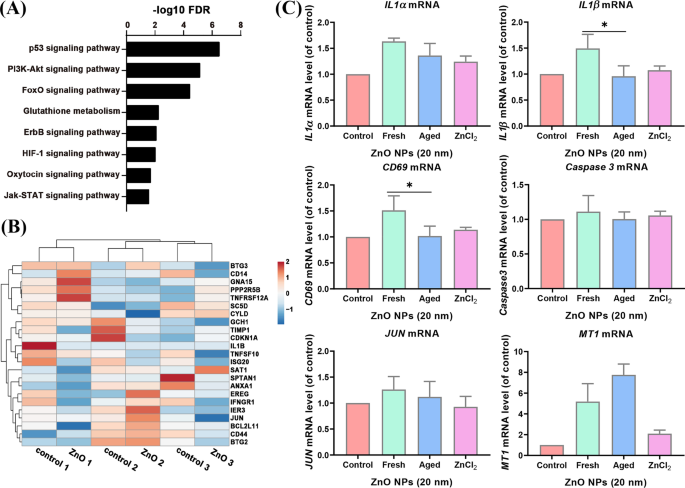
A via de apoptose não foi enriquecida em dados de RNA-seq de idosos tratados com ZnO NP A L células. ( A ) A análise de ontologia genética de vias enriquecidas de envelhecidos tratados com ZnO NP A L células. ( B ) O mapa de calor da expressão de genes apoptóticos de envelhecidos ZnO NP-tratados A L células e grupo controle. ( C ) A expressão de genes apoptóticos selecionados e genes de controle ( MT1 ) em A tratado com ZnO NP fresco e envelhecido L células
NPs de ZnO frescos, mas não envelhecidos, aumentaram o nível de expressão da proteína caspase 3 ativada
Detecção de expressão gênica de Caspase 3 por si só não pode indicar diretamente a ativação da via de apoptose. Para uma análise mais aprofundada se o tratamento com ZnO NPs poderia alterar o nível de proteínas apoptóticas, a expressão da proteína Caspase 3 clivada, um biomarcador comumente usado para indicar a ativação da apoptose celular [27], foi examinada por análise de Western blotting. Como mostrado na Fig. 6, em comparação com o grupo de controle, o tratamento com ZnO NPs (20 nm) fresco aumentou o nível celular da proteína Caspase 3 clivada em 1,31 ± 0,023 vezes, o que foi significativamente maior do que o de 20 nm ZnO NPs- grupo tratado (1,12 ± 0,039 vezes). Quando o tamanho de partícula de NPs de ZnO frescos foi aumentado para 90-200 nm, a expressão da proteína Caspase 3 clivada induzida por NPs frescos foi aumentada em 1,46 ± 0,078 vezes, significativamente maior do que a de NPs envelhecidos (1,07 ± 0,075 vezes) . Esses dados ilustraram ainda a maior potência de NPs de ZnO frescos na indução da apoptose celular, em comparação com suas contrapartes envelhecidas.
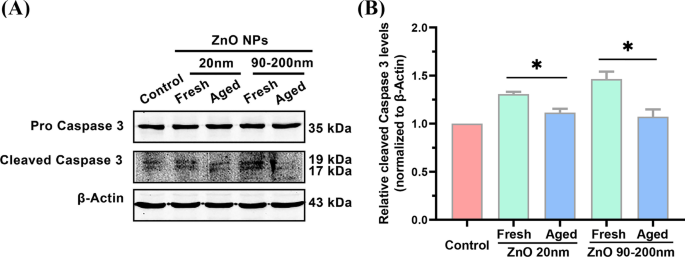
Níveis apoptóticos em A L células incubadas com ZnO NPs frescos e envelhecidos (20 e 90–200 nm). Análise de Western Blotting ( A ) e quantificação ( B ) dos níveis de proteína Caspase 3 clivada quando as células foram incubadas com 12 μg / mL de NPs de ZnO frescos e envelhecidos (20 e 90–200 nm) por 72 h. Os dados foram baseados em ≥ 3 experimentos independentes e expressos como média ± DP, * p <0,05
Discussão
Foi relatado que NPs de ZnO sofreram transformação físico-química em Zn 5 (CO 3 ) 2 (OH) 6 com o lançamento de Zn 2+ durante o processo natural de envelhecimento [23, 28]. No entanto, a citotoxicidade induzida pelos NPs de ZnO transformados (envelhecidos) e os mecanismos subjacentes permanecem obscuros. Aqui, para desvendar o mecanismo de citotoxicidade diversa entre ZnO NPs frescos e envelhecidos, foram realizadas análises de sequenciamento de RNA e teste de RT-PCR. Além disso, Western blotting foi aplicado para examinar o nível de proteína da Caspase 3, o executor chave na apoptose celular.
Nossos dados mostraram que NPs de ZnO envelhecidos induziram muito menos citotoxicidade do que NPs de ZnO frescos em A L células. O LC 100 de ambos os NPs de ZnO frescos (90–200 nm e 20 nm) em nosso presente estudo foi inferior a 15 μg / mL (Fig. 3), o que foi consistente com os achados anteriores de que o LC 100 de NPs de ZnO com 19-36 nm para células NIH-3T3 ou MSTO é cerca de 15 μg / mL [29]. Confirmamos que as transformações ambientais das propriedades físico-químicas em NPs podem alterar dramaticamente sua toxicidade. Foi relatado que o processo de sulfetação de NPs de ZnO altera sua carga, hidrofobicidade e estado de agregação, resultando na adsorção de NPs em estado de sulfeto na saliva humana, suor e fluido de lavagem broncoalveolar. E a proteína adsorvida pelos NPs de ZnO formou uma coroa protéica especial, que afetou seu efeito biológico [19]. Os fosfatos amplamente presentes em soluções fisiológicas (como a saliva) podem converter ZnO NPs em ZnHPO metaestável 4 e Zn 3 (PO 4 ) 2 em cerca de 5–10 he mostrou citotoxicidade para células epiteliais do trato digestivo [20]. Ivask et al. comprovou a ocorrência de transformação completa de ZnO NPs (≤ 3 μg / mL) no sistema de exposição in vitro para linfócitos T humanos (37 ° C, meio de cultura celular RPMI1640 contendo 10% de FBS por 24 h) usando absorção de raios-X de radiação síncrotron espectroscopia de estrutura próxima à borda (XANES). O espectro e a citotoxicidade dos produtos de transformação eram consistentes com os de ZnSO 4 [21]. Nossos resultados revelaram a toxicidade dependente da dose e do tempo de ZnCl 2 para A L células, onde sua citotoxicidade é muito menor do que as NPs de ZnO frescas e envelhecidas (Fig. 3). A observação explica ainda a descoberta de que a citotoxicidade do ZnO NP fresco não é totalmente atribuída ao seu Zn 2+ liberado [30].
Nosso estudo anterior também mostrou que os NPs de ZnO envelhecidos exibem uma maior potência na eliciação de ROS (espécies reativas de oxigênio), bem como uma capacidade atenuada em matar células em comparação com os NPs de ZnO frescos [23]. Raciocinamos que a menor citotoxicidade induzida por NPs de ZnO envelhecidos poderia ser mais tolerável em células de mamíferos. O presente estudo de dados de sequenciamento de RNA ilustrou que os genes apoptóticos foram regulados positivamente em células tratadas com ZnO NP frescas, onde foram muito menos afetados em grupos tratados com NP envelhecidos. IL1α e IL1β são membros da família de citocinas da interleucina 1. A liberação de IL1α e IL1β ativa a apoptose parcialmente dependente da Caspase 8 [31]. CD69 codifica um membro da superfamília de lectina dependente de cálcio de receptores transmembrana de tipo II. A expressão aumentada de CD69 foi associada a uma expressão aumentada do marcador de apoptose anexina V e CD95 (Fas) [32]. JUN é uma subunidade do fator de transcrição AP-1. A atividade aumentada da JUN cliva proteoliticamente a alfa-fodrin, um substrato da enzima conversora de interleucina 1beta (ICE), e da família CED-3 de proteases de cisteína, que ainda causa morte celular programada [33]. O aumento da expressão desses genes apoptóticos revelou que NPs recentes desencadeiam a apoptose de várias maneiras diferentes. Após a elevação dessas expressões gênicas apoptóticas, os processos de apoptose são eventualmente executados por proteínas apoptóticas (Fig. 7). Caspase 3 é a protease central para vários cenários apoptóticos; a clivagem desta proteína é necessária para ativar as vias apoptóticas extrínsecas e intrínsecas [34, 35]. Therefore, detection of cleaved caspase 3 is a common method for identifying apoptosis induced by a wide variety of apoptotic signals [36]. Our Western blotting data revealed that, for both 20 nm and 90–200 nm ZnO NPs, sublethal exposure did not alter the level of Pro caspase 3 in all treatment groups. In contrast, cleaved Caspase 3 was significantly elevated by fresh NPs treatment, where aged NPs showed few (if any) effects on the level of cleaved caspase 3 (Fig. 6). Combined with RNA expression analysis, our results clearly elucidated the higher potency of fresh ZnO NPs in inducing cell apoptosis.

Model for Fresh ZnO NPs but not aged ZnO NPs induces Caspase 8- and Caspase 3-dependent apoptosis. The increased expression of apoptotic gene CD69 activates Fas and apoptosis annexin V expression in fresh ZnO NP-exposed mammalian cells. The increased expression of apoptotic gene IL1α and IL1β partially activates Caspase 8-dependent apoptosis. It further causes activation of Caspase 3 and induces apoptosis. All these changes in mRNA and protein level were not detectable in aged ZnO NPs-exposed mammalian cells
Conclusões
In the present study, the natural physicochemical transformation of ZnO NPs in ultrapure water was confirmed, and variations in cytotoxicity induced by fresh &aged NPs were investigated. We focused on RNA sequencing data from our aged ZnO NP-treated A L cells and that of fresh NPs from database. We compared those signaling pathway specifically enriched in aged NP-treated group, which are different from that of fresh NP- or ZnCl2 -treated groups. Our data indicated that the lower cytotoxicity of aged ZnO NPs is closely related to its attenuated ability in inducing apoptosis, while the transcriptional regulation of the multiple pathways activated by NPs promotes the establishment of cellular homeostasis in mammalian cells.
Disponibilidade de dados e materiais
Não aplicável.
Abreviações
- NPs:
-
Nanopowders
- ZnO:
-
Óxido de zinco
- Zn5 (CO 3 ) 2 (OH) 6 :
-
Hydrozincite
- Zn (OH)2 :
-
Zinc hydroxide
- ZnCl2 :
-
Zinc chloride
- ZnSO4 :
-
Zinc sulfide
- ZnHPO4 :
-
Zinc hydrogen phosphate
- Zn3 (PO 4 ) 2 :
-
Zinc phosphate
- A L cells:
-
Human–hamster hybrid cells
- CHO cells:
-
Chinese hamster ovary cells
- Jurket cells:
-
Peripheral blood T lymphocyte cells
- HMDM cells:
-
Human monocyte-derived macrophages
- NIH-3T3cells:
-
Mouse embryonic cells
- MSTO cells:
-
Human lung cancer cells
- RPMI1640:
-
Roswell Park Memorial Institute 1640
- ICE:
-
Interleukin 1beta-converting enzyme
- CED-3:
-
Caenorhabditis elegans death gene
- IL1α:
-
Interleukin 1alpha
- IL1β:
-
Interleukin 1beta
- mRNA:
-
Messenger ribonucleic acid
- cDNA:
-
Complementary deoxyribonucleic acid
- FBS:
-
Soro fetal bovino
- TEM:
-
Microscopia eletrônica de transmissão
- XRD:
-
Difração de raios X
- RT-PCR:
-
Real-time polymerase chain reaction
- CPI:
-
The Nanotechnology Consumer Product Inventory
- XANES:
-
Synchrotron radiation X-ray absorption near-edge structure spectroscopy
- RIPA:
-
Radio immunoprecipitation assay
- SDS-PAGE:
-
Polyacrylamide gel electrophoresis
- PVDF:
-
Fluoreto de polivinilideno
- ECL:
-
Enhanced chemiluminescence
- CCK-8:
-
Kit de contagem de células-8
Defeito e doping Penta-grafeno projetado para catálise da reação de evolução de hidrogênio
Troca de limite do seletor Ag-Ga2Te3 com alta resistência para aplicações em matrizes de ponto cruzado
Nanomateriais
- Nanopartículas de FePO4 biocompatíveis:entrega de drogas, estabilização de RNA e atividade funcional
- Efeitos da Espessura de Bicamada nas Propriedades Morfológicas, Óticas e Elétricas de Nanolaminados de Al2O3 / ZnO
- Ajustando as morfologias de superfície e propriedades dos filmes ZnO pelo projeto da camada interfacial
- Adepto verde na síntese e estabilização de nanopartículas de cobre:atividades catalíticas, antibacterianas, citotoxicidade e antioxidantes
- Fabricação e caracterização de Nano-Clips de ZnO pelo processo mediado por poliol
- Uma análise comparativa in vivo de nanopartículas de óxido de cobre e zinco biossintetizadas por vias de administração intraperitoneal e intravenosa em ratos
- Síntese Verde de Nanopartículas de Metal e Óxido de Metal e Seu Efeito na Alga Unicelular Chlamydomonas reinhardtii
- Propriedades das nanopartículas de óxido de zinco e sua atividade contra micróbios
- Uma abordagem fácil para a preparação de óxido de zinco de tamanho nanométrico em água / glicerol com fontes de zinco extremamente concentradas
- O papel da robótica e da automação na indústria 4.0