


SPIO melhora a apresentação cruzada e migração de DCs e SPIO aniônico influencia os efeitos nanoadjuvantes relacionados à interleucina-1β
Resumo
Nanopartículas de óxido de ferro superparamagnético (SPIO) foram sintetizadas e exploradas para uso como transportadores de vários nanoadjuvantes via carregamento em células dendríticas (DCs). Em nosso estudo, nanopartículas homogêneas e superparamagnéticas são suscetíveis à internalização por DCs e as DCs pulsadas com SPIO mostraram excelente biocompatibilidade e capacidade de apresentação cruzada de ovalbumina (OVA). Aqui, descobrimos que DCs carregados com SPIO podem promover a maturação e migração de DCs in vitro . Foram preparados SPIO revestidos com 3-aminopropiltrimetoxissilano (APTS) e ácido meso-2,3-dimercaptosuccínico (DMSA), que apresentam cargas positivas e negativas, respectivamente. Nosso objetivo foi investigar se a carga superficial de SPIO pode afetar a apresentação cruzada de antígenos das DCs. Além disso, a formação de interleucina-1β (IL-1β) foi examinada após o tratamento com SPIO de carga oposta para identificar o mecanismo de nanoadjuvantes. Em conclusão, nossos resultados sugerem que SPIO são biocompatíveis e podem induzir a migração de DCs para linfonodos secundários. SPIO revestido com APTS (SPIO / A + ) exibiram excelentes potenciais adjuvantes para a promoção de apresentação cruzada de antígenos e ativação de células T e ultrapassaram o de nanopartículas revestidas com DMSA (SPIO / D - ) Esse processo pode estar relacionado à secreção de IL-1β. Nosso estudo fornece insights sobre a modificação preditiva de nanoadjuvantes, que serão valiosos no projeto de vacinas DC e podem levar à criação de novos adjuvantes para aplicações em vacinas para humanos.
Histórico
Os adjuvantes têm sido aplicados extensivamente para uso clínico e experimental e há muito tempo são considerados agentes imunoestimulantes, depósitos passivos ou veículos capazes de promover as respostas imunes necessárias [1, 2]. Nas últimas décadas, muitas classes diversas de compostos têm sido usadas como adjuvantes, incluindo nanopartículas, produtos microbianos, emulsões, citocinas, polímeros e lipossomas [3,4,5]. A evolução das nanopartículas como adjuvantes imunológicos (nanoadjuvantes) representa uma área notável de entrega de antígenos com base na ampliação das respostas imunológicas. Dentre todos os tipos de nanopartículas, as nanopartículas de óxido de ferro superparamagnéticas (SPIO) têm grande biocompatibilidade, arquitetura de superfície adequada e conjugação de ligante flexível [6]. Essas propriedades os tornam aplicáveis em muitas áreas biomédicas diferentes, como imagem por ressonância magnética (MRI), administração de drogas direcionadas e terapia de hipertermia [7, 8].
As células dendríticas (DCs), as principais células apresentadoras de antígenos profissionais (APCs), desempenham um papel crítico na imunidade adaptativa mediada por células, na qual a memória imunológica é gerada após uma resposta primária a um antígeno específico, e essa memória leva a uma resposta aprimorada a encontros subsequentes com aquele antígeno [9, 10]. Além disso, as DCs podem promover a apresentação de antígenos exógenos pelo complexo principal de histocompatibilidade classe I (MHC-I), um processo denominado apresentação cruzada, e então ativar os linfócitos T citotóxicos (CTLs) [11]. Antígenos solúveis destinados à apresentação cruzada são internalizados por endocitose mediada por receptor e, em seguida, transferidos para o citoplasma para degradação do proteassoma e carregamento de peptídeo. As vacinas convencionais de DC levam a uma resposta imunológica moderada devido ao transporte relativamente insatisfatório de antígenos solúveis. Portanto, os nanoadjuvantes têm sido explorados como transportadores de antígenos solúveis para aumentar a apresentação cruzada em DCs em muitos estudos de pesquisa [12,13,14].
Os materiais de revestimento desempenham papéis críticos na estabilização e subsequente funcionalização de suspensões aquosas de SPIO [15]. Em estudos anteriores, SPIO carregados positivamente foram mostrados para facilitar a capacidade de apresentação cruzada de DCs para aumentar a resposta imune, e eles superaram o efeito de suas contrapartes com carga oposta [16, 17]. O mecanismo pelo qual SPIO com carga diferente pode afetar a apresentação cruzada de antígenos de DCs não foi esclarecido. A interleucina-1β (IL-1β), uma citocina pró-inflamatória prototípica que participa da imunidade inata, pode ser secretada por células imunes, como DCs, ao detectar padrões moleculares associados a patógenos (PAMPs) e padrões moleculares associados a danos (DAMPs) [ 18]. A produção de IL-1β é estritamente mediada por inflamassomas, especialmente NLRP3 (proteína 3 contendo os domínios NACHT, LRR e PYD). A ativação de NLRP3 induz a produção de caspase-1, que então cliva a pró-IL-1β inativa na forma ativa IL-1β [19]. Certas nanopartículas inorgânicas, como sílica, nanotubos de carbono de parede dupla e dióxido de titânio, podem induzir a formação de inflamassoma [20,21,22]. Estudos anteriores relataram uma correlação entre a carga superficial das nanopartículas de magnetita e sua eficiência celular [23]. Aqui, especulamos que diferentes cargas de superfície em DCs carregadas com SPIO também podem influenciar a secreção de IL-1β, e nosso objetivo é investigar a relação entre essas cargas de superfície e a função de apresentação cruzada de DCs.
Métodos e materiais
Preparação do SPIO
Para preparar SPIO, um método fácil de coprecipitação foi empregado, conforme relatado anteriormente [24]. Em resumo, uma solução mista de FeCl 3 e FeSO 4 (razão molar Fe 3+ :Fe 2+ =2:1) foi preparado sob atmosfera de nitrogênio e energicamente agitado a 37 ° C por 30 min. Um precipitado de cor preta de Fe 3 O 4 nanopartículas foram formadas e imediatamente lavadas cinco vezes com água destilada usando separação magnética. The Fe 3 O 4 foi então disperso em água destilada a uma concentração de 3 mg / mL a um pH de 3. Por fim, a suspensão foi aerada (com ar) a 95 ° C e o SPIO marrom foi separado.
SPIO revestido com DMSA (SPIO / D - ) e SPIO revestido com APTS (SPIO / A + ) foram preparados revestindo SPIO com ácido meso-2,3-dimercaptosuccínico (DMSA) e 3-aminopropiltrimetoxissilano (APTS), respectivamente. Para SPIO / D - , uma solução aquosa de DMSA em uma razão molar de 1:40 foi adicionada a 100 mL de solução de SPIO. Após uma reação de 4 h com agitação contínua, o SPIO / D - foi separado a uma taxa de 500 rpm a 50 ° C. Para SPIO / A + , APTS foi adicionado à solução de SPIO a uma razão molar de 0,2:1 com agitação vigorosa durante 5 h. Em seguida, o precipitado foi dissolvido com um ímã permanente e lavado com água deionizada, e a solução foi tratada por ondas ultrassônicas. A solução resultante foi lavada repetidamente com água, e o SPIO / A + foram finalmente secos em pó a 37 ° C sob vácuo.
Ratos
Camundongos C57BL / 6 e proteína fluorescente verde aprimorada (EGFP) -transgênica C57BL / 6 camundongos foram adquiridos do Model Animal Research Center da Universidade de Nanjing e alojados em condições específicas livres de patógenos (SPF) no Laboratório Central da Universidade de Nanjing. Todas as experiências com animais foram realizadas de acordo com os protocolos aprovados pelo Animal Care and Use Committee da Faculdade de Medicina da Universidade de Nanjing, China.
Cultura de células
As DCs murinas foram geradas a partir da medula óssea de camundongos, conforme descrito anteriormente [25]. Em resumo, os monócitos da medula óssea de camundongos C57BL / 6 de 8 semanas de idade foram separados de seus fêmures e tíbias. Posteriormente, as células foram cultivadas em RPMI-1640 (Gibco, Thermo Fisher Scientific, Waltham, MA, EUA) junto com 10% de soro fetal bovino (FBS), 10 ng / mL de fator estimulador de colônia de granulócito-macrófago murino (GM-CSF; Gibco, EUA), e 1 ng / mL de interleucina-4 murina (IL-4, PeproTech, Rocky Hill, NJ, EUA). O meio de cultura foi substituído por meio fresco a cada 2 dias. As DCs imaturas foram geralmente coletadas no dia 6. As EGFP-DCs foram derivadas de camundongos C57BL / 6 transgênicos EGFP de acordo com o método mencionado acima.
Células mononucleares de sangue periférico humano descongeladas (PBMCs) foram repousadas durante a noite em meio completo, isto é, X-VIVOTM 15 (Lonza Group Ltd., Basel, Suíça) =1:1. Após centrifugação em gradiente de densidade, os PBMCs foram suspensos em meio por 4 h. DCs humanas das células aderentes foram cultivadas a 37 ° C em meio contendo GM-CSF humano (100 ng / mL; R&D Systems, Minneapolis, MN, EUA) e IL-4 humana (10 ng / mL; PeproTech, Rocky Hill, NJ, EUA). Metade do meio foi trocado nos dias 2 e 4. DCs humanos imaturos foram colhidos no dia 5. Células T específicas do vírus Epstein-Barr do citomegalovírus e do vírus influenza (CEF) foram derivados de células suspensas, que foram cultivadas em meio CMX contendo MHC -I restringiu o peptídeo CEF (20 ng / mL, Panatecs, Heilbronn, Germany, PA-CEF-002) por aproximadamente 48 h. As células T específicas para CEF foram expandidas por 1000 U / mL de IL-2 humana (Peprotech, Rocky Hill, NJ, EUA, 200-02) em meio CMX por 12 dias. Para IL-2, o meio foi substituído por meio fresco a cada 3 dias.
Caracterização
A morfologia e o tamanho do SPIO foram determinados usando um microscópio eletrônico de transmissão (TEM, Advanced Microscopy Techniques, Danvers, MA). Um padrão de difração de raios-X (XRD) foi usado para identificar os espectros do catalisador. O ensaio de potencial zeta também foi medido para determinar as cargas de superfície das nanopartículas revestidas com diferentes polímeros. O SPIO / A + e SPIO / D - nanopartículas foram preparadas com valores de pH variando de 3 a 8. As medições do potencial zeta foram feitas com um analisador de potencial zetasizer Nano ZS90 (Malvern, Reino Unido). As propriedades magnéticas das nanopartículas foram analisadas a 37 ° C usando um magnetômetro de amostra vibrante (Lakeshore 7407).
Ensaio de CPRG
O B 3 Linha de células Z T (um CD8 + Hibridoma de células T) pode expressar o gene LacZ quando seu receptor de células T envolve um epítopo de ovalbumina (OVA) 258–265 na presença da molécula H-2Kb MHC-I. DCs (2 × 10 4 ) e OVA (100 μg / mL, Sigma-Aldrich) foram cultivados com SPIO, SPIO / A + ou SPIO / D - a 37 ° C. Seis horas depois, os DCs foram co-cultivados com B 3 Z (2 × 10 5 ) durante a noite. Um ensaio de clorofenol red-β-galactosidase (CPRG, Sigma-Aldrich, EUA) foi conduzido para determinar a produção de β-galactosidase do B 3 Células Z. Neste ensaio, a densidade óptica (OD) a 595 nm indica a capacidade de apresentação cruzada do antígeno das DCs.
Ensaio de apoptose de células
A classificação de células ativadas por fluorescência (FCS) foi empregada para explorar os efeitos de diferentes concentrações de SPIO na viabilidade das DCs. Em resumo, DCs imaturas foram incubadas com SPIO marcado com Anexina V e PI (Biouniquer, CHN), e a expressão de Anexina V e PI nas DCs foram examinadas via FCS.
Imagem de intensidade de fluorescência in vivo
Para explorar a migração de DCs in vivo, TNF-α (60 ng / camundongo) foi pré-injetado nas patas de ambas as patas traseiras ( n =8). Após 24 h, EGFP-DCs marcados com SPIO (2 × 10 6 ) em 40 μL de solução salina tamponada com fosfato (PBS) foram injetados na planta da pata esquerda de camundongos C57BL / 6, e o mesmo número de EGFP-DCs não marcados foi injetado no lado direito. Para examinar o nível de EGFP-DCs migrando para os nódulos linfáticos, um sistema de imagem Maestro (CRi, Woburn, MA, EUA) foi empregado. Os linfonodos dissecados foram observados usando filtros de excitação de 484 nm e filtros de emissão de 507 nm. As imagens fluorescentes exibindo fluorescência verde foram então analisadas com o software Living Image (v 2.50; Caliper Corporation, Newton, MA, EUA). Análises de microscopia confocal foram utilizadas para determinar a imunohistoquímica. Os gânglios linfáticos congelados foram cortados em seções de 5 μm de espessura e, em seguida, fixados, e as seções foram incubadas com anticorpo de proteína fluorescente verde (GFP) (Invitrogen), com anticorpo Alexa Fluor 488 nm de cabra (Invitrogen) usado como anticorpo secundário. Um microscópio confocal de varredura a laser (Fluoview, Fv10i; Olympus) foi usado para observar as amostras.
Ensaio de apresentação cruzada de antígeno DC humano in vitro
As DCs humanas cultivadas (2 × 10 4 ) foram semeados em uma placa de fundo redondo de 96 poços para a qual SPIO / A + e SPIO / D - nanopartículas (100 μg / mL) combinadas com proteína pp65 do citomegalovírus (CMV) (1 μg / mL, MACS) foram adicionadas e a proteína pp65 de CMV e o peptídeo CEF (1 μg / mL, peptídeos Think) foram usados separadamente como controles. Após 6 h, células T específicas de CEF (2 × 10 5 ) foram adicionados às DCs humanas por 12 h. Brefeldin A (10 μg / ml, Sigma-Aldrich, EUA) foi mantida no meio por mais 6 h. Após estimulação e ativação, células T específicas de CEF foram reunidas e coradas com anticorpos humanos contra CD3, CD4, CD8 e IFN-γ (Invitrogen, EUA). Após a coloração, as amostras foram analisadas por um citômetro de fluxo LSR II personalizado (BD, Franklin Lakes, NJ, EUA). Os dados coletados foram analisados pelo software FlowJo (Tree Star, Ashland, OR, EUA).
Ensaio de imunoabsorção enzimática
Um kit de ensaio de imunoabsorção enzimática (ELISA) Ready SET-Go de IL-1β de camundongo (eBioscience, EUA) e um kit ELISA de IL-1β humana (Thermo Fisher Scientific, Waltham, MA, EUA) foram empregados para determinar a secreção de IL -1β por DCs. Uma placa ELISA (Costar, EUA) revestida com 100 μL / poço de IFN-γ foi usada para capturar anticorpos durante a noite a 4 ° C e bloqueada com tampão ELISA. Amostras e padrões foram então adicionados aos poços e incubados por 2 h a 37 ° C. Foi detectado IFN-γ biotinilado. As amostras foram medidas em um leitor de microplacas com ajuste de DO de 450 nm (BioTek, EUA).
Análise estatística
Os dados foram analisados por meio do Statistical Package for Social Science (SPSS 13.0, Chicago, IL, EUA). Os resultados foram apresentados como as médias ± DP, e as diferenças entre os grupos de controle e teste foram avaliadas por uma análise de variância unilateral, t de Student bicaudal testes e análise de variância de fator duplo. Diferenças em * P <0,05 foram considerados estatisticamente significativos.
Resultados
Caracterização de SPIO
A morfologia e o tamanho do SPIO sintetizado foram observados via TEM. A imagem TEM mostrou que SPIO tinha um tamanho médio de 8,7 nm e uma forma esférica (Fig. 1a). A análise de XRD mostrou seis picos que correspondiam distintamente ao γ-Fe 2 padrão O 3 reflexões (Fig. 1b). Os resultados do magnetômetro vibratório demonstraram que o SPIO obtido possuía comportamento superparamagnético, com uma magnetização de saturação de 60,4 emu / g melhor do que Fe 3 O 4 (Fig. 1c). O gráfico DLS mostrou que a distribuição de tamanho do SPIO é de 22 nm em solução (Fig. 1d). Para confirmar que as DCs continham SPIO, a coloração com azul da Prússia foi realizada para verificar se as DCs continham ferro (Fig. 1e).
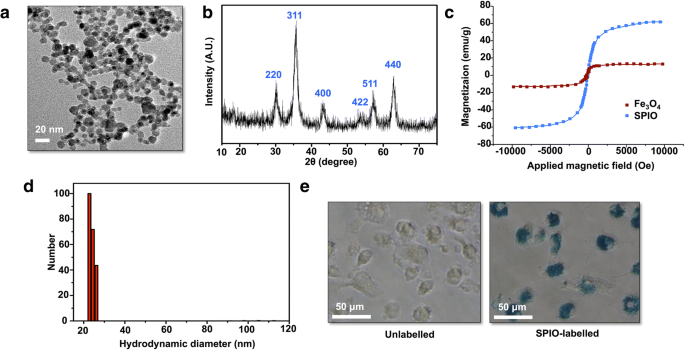
Caracterização do SPIO. a Imagem TEM do SPIO obtido. b Padrão de XRD do catalisador de nanopartículas indicando que o material é γ-Fe 2 O 3 . c Curvas de magnetização do SPIO obtido e Fe 3 O 4 nanopartículas. d Diâmetro hidrodinâmico de SPIO. e Morfologia das DCs marcadas com 50 μg / mL SPIO após 12 h de incubação:DCs não marcadas e DCs marcadas com azul da Prússia
Apresentação cruzada de CDs ativada por SPIO
Para estudar ainda mais o efeito das DCs marcadas com SPIO na ativação de células T em um sistema murino, o nível de B 3 A ativação de células Z T foi determinada examinando a produção de β-galactosidase por ensaio de CPRG. Uma concentração fixa de 100 μg / mL de OVA e cinco proporções de dose de SPIO (1, 5, 10, 25 e 50 μg / mL após 6 h) foram adotadas neste estudo. Conforme a concentração de SPIO aumentou, o grau de ativação de B 3 As células Z aumentaram gradualmente e alcançaram estabilidade em 25 μg / mL (Fig. 2a). Para investigar se a viabilidade das DCs marcadas com as várias concentrações de SPIO foi influenciada, as DCs e as DCs marcadas com SPIO foram analisadas via FCS após a coloração com Anexina V e PI. Os resultados indicaram que a porcentagem total de DCs de Anexina V / PI em concentrações de SPIO de 10, 25 e 50 μg / mL não diferiram significativamente, enquanto a porcentagem de células apoptóticas aumentou após o carregamento com SPIO de 100 μg / mL (Fig. 2b ) As moléculas coestimulatórias de superfície das DCs marcadas com várias concentrações de SPIO foram observadas por FCS. A expressão de CD80 e CD86 teve um aumento detectável de 25 μg / mL em comparação com aqueles sem marcação SPIO (Fig. 2c). DCs marcados com 25 μg / mL SPIO não exibiram alterações na apoptose celular em pontos de tempo diferentes (Fig. 2d). Usamos 25 μg / mL nos seguintes experimentos.
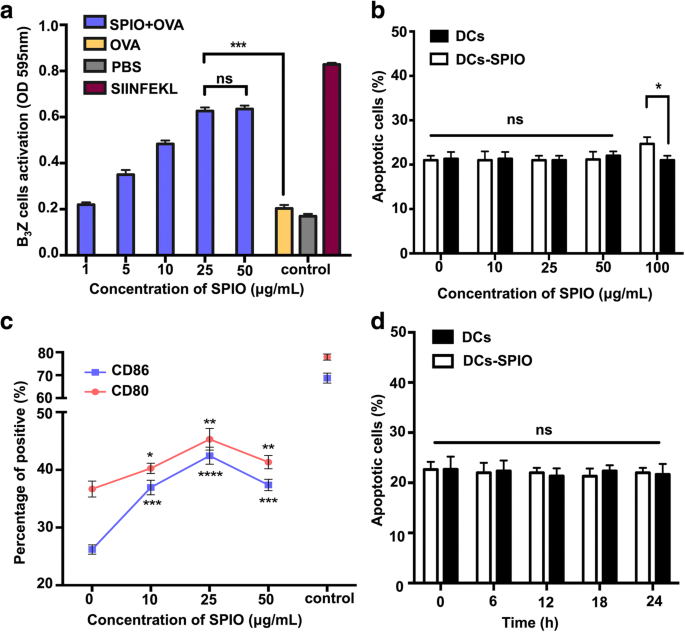
Influências da apresentação cruzada de DC e biocompatibilidade após rotulagem com SPIO. a Apresentação cruzada de DCs aprimorada em diferentes concentrações de SPIO. b DCs apoptóticas e DCs marcadas com SPIO examinadas por FCS em diferentes concentrações (10, 25, 50 e 100 μg / mL). c Fenótipos dos DCs, incluindo CD11c + CD80 + e CD11c + CD86 + após a marcação com SPIO (10, 25, 50 μg / mL). DCs estimuladas com LPS (1 μg / mL) foram usadas como controles positivos. d DCs apoptóticas carregadas com 25 μg / mL SPIO examinados por FCS em diferentes pontos de tempo (6, 12, 18 e 24 h)
Rotulando EGFP-DCs com sinais de EGFP aprimorados SPIO em linfonodos secundários
EGFP-DCs foram derivados com sucesso de camundongos transgênicos EGFP in vitro, e imagens de microscopia de fluorescência confocal mostraram que quase todos os EGFP-DCs exibiram fluorescência verde (Fig. 3a). Para investigar se a fluorescência verde das EGFP-DCs poderia ser afetada por SPIO, o FCS foi conduzido. Os resultados mostraram que a expressão da fluorescência EGFP não foi enfraquecida após a marcação SPIO (Fig. 3b, c). Em seguida, os 25 μg / mL de EGFP-DCs marcados com SPIO foram injetados nas patas traseiras do lado direito de camundongos C57BL / 6 e EGFP-DCs não marcados foram injetados no lado oposto. Os sinais de EGFP foram medidos no linfonodo poplíteo (PLN) e no linfonodo inguinal (ILN), que são os linfonodos sentinela e os linfonodos secundários, respectivamente. Os resultados mostraram que a migração de EGFP-DCs marcados com SPIO e EGFP-DCs não marcados nos linfonodos sentinela atingiu um pico no dia 7. Diferenças significativas no sinal de EGFP não foram observadas entre os dois grupos, enquanto uma redução significativa foi detectada no grupo PLN no dia 14. Os sinais de EGFP detectados no grupo ILN foram consistentes com os do grupo EGFP-DC marcado com SPIO no 4º dia e no 7º dia, o que indicou que as DCs marcadas com SPIO migraram para a linfa secundária nós (Fig. 3d-f).
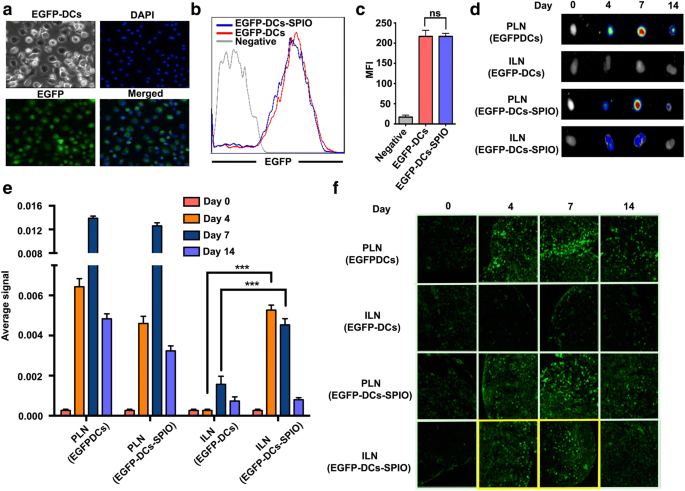
Migração e localização de EGFP-DCs após rotulagem com SPIO. a Microscopia confocal de varredura a laser de EGFP-DCs marcados com 25 μg / mL de nanopartículas de SPIO após 12 h de incubação. b , c Intensidade de fluorescência das EGFP-DCs marcadas com SPIO após 12 h. Imagem óptica in vitro de linfonodos de drenagem após a retrotransfusão de EGFP-DCs marcados com SPIO em dias diferentes. O TNF-α foi primeiro injetado no tapete do rato com antecedência para promover a migração de EGFP-DCs. d - e Imagem óptica in vitro e análise de intensidade de sinal de nódulos linfáticos em dias diferentes após a injeção de EGFP-DCs com ou sem SPIO. f Células EGFP positivas de linfonodos de drenagem foram detectadas por microscopia confocal a laser
Apresentação cruzada de DCs murinos afetados por SPIO modificados de forma diferente
Os SPIO foram revestidos com APTS ou DMSA. Para verificar a carga superficial do SPIO, medimos a característica do potencial zeta. Os potenciais zeta do SPIO / A + e SPIO foram positivos, enquanto o potencial zeta do SPIO / D - mostrou uma carga negativa em solução quando o valor de pH era 7 (Fig. 4a). A ultraestrutura das DCs marcadas com SPIO revestidas com polímeros de carga diferente foi observada via TEM. As DCs tratadas com SPIO pareciam elétron-densas em comparação com as células não tratadas, e numerosas SPIO foram agrupadas no citoplasma. O SPIO / A + foram engolfados por endossomos no citoplasma, enquanto uma quantidade maior de SPIO / D - foram encontrados no citoplasma das DCs, e quase todas essas nanopartículas foram circundadas por estruturas de membrana de várias camadas que se assemelhavam a lisossomas (Fig. 4b). Nós examinamos se o SPIO com carga oposta poderia desencadear diferentes níveis de IL-1β. De acordo com nossos resultados, DCs carregados com SPIO / A + e SPIO / D - induziu uma secreção dependente da dose de IL-1β. Independentemente da concentração, descobrimos que SPIO / D - induziu níveis significativamente mais elevados de IL-1β do que SPIO / A + (Fig. 4c). Devido à correlação óbvia entre IL-1β e a via de TLR3, para estudar mais o efeito de IL-1β na ativação de células T, as DCs foram derivadas de camundongos knockout para TLR3 e co-cultivadas com SPIO / A + , SPIO / D - e OVA. Os DCs carregados com SPIO / A + + OVA poderia efetivamente ativar B 3 Células Z T, o que significa que a molécula TLR3 nas DCs foi definitivamente necessária para apresentação cruzada (Fig. 4d).
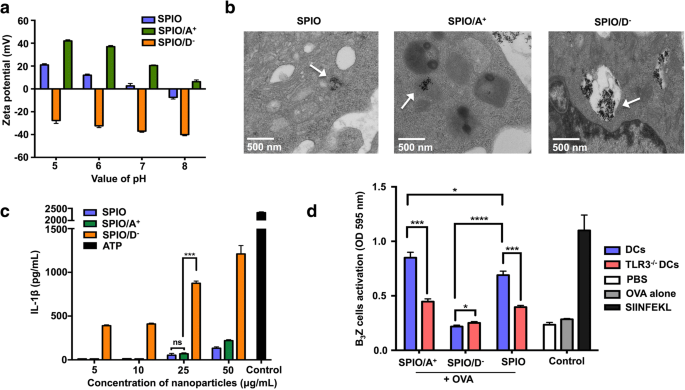
SPIO revestido com cargas diferentes afetou a apresentação cruzada de DC e a secreção de IL-1β. a O potencial zeta dependente do pH de SPIO revestido com moléculas com cargas diferentes. b Localizações de nanopartículas com diferentes cargas em DCs sob TEM. c IL-1β induzida por SPIO revestida com moléculas com cargas diferentes. d Apresentação cruzada de SPIO / A + + OVA e SPIO / D - + OVA por DCs através da via TLR3
SPIO modificado com revestimentos de carga oposta promoveu apresentação cruzada de antígeno por DCs humanos e foi afetado por IL-1β
Para explorar a relação entre IL-1β e apresentação cruzada, selecionamos o inibidor de caspase-1 YVAD e descobrimos que ele poderia inibir significativamente a secreção de IL-1β de DCs humanas após o pré-tratamento com 50 μM de lipopolissacarídeo (LPS) por 3 h (Fig. . 5a). Além disso, descobrimos que YVAD pode inibir a secreção de IL-1β de DCs humanas causada por SPIO / A + e SPIO / D - (Fig. 5b). Em seguida, investigamos se as DCs humanas poderiam ser usadas como APCs eficazes e apresentar CMV pp65 de forma cruzada com células T específicas de CEF. Para medir a apresentação cruzada do antígeno, a coloração intracelular (ICS) foi introduzida para determinar a porcentagem de CD3 específico do antígeno + CD8 + IFN-γ + e CD3 + CD4 + IFN-γ + Células T por FCS. DCs carregados com SPIO / A + combinado com a proteína CMV pp65 induziu mais CD3 + CD8 + IFN-γ + e CD3 + CD4 + IFN-γ + Células T do que DCs carregadas com SPIO / D - . Nossos dados mostraram que o inibidor da caspase-1 YVAD aumentou significativamente as respostas das células T induzidas pelo SPIO / D - + Proteína pp65 do CMV, ao passo que inibiu parcialmente as respostas das células T induzidas pelo SPIO / A + + Proteína pp65 de CMV (Fig. 5c, d). Esses resultados indicaram que um nível moderado de ativação de IL-1β é necessário para apresentação cruzada eficiente e que um alto nível de ativação de IL-1β suprime a apresentação cruzada em DCs.
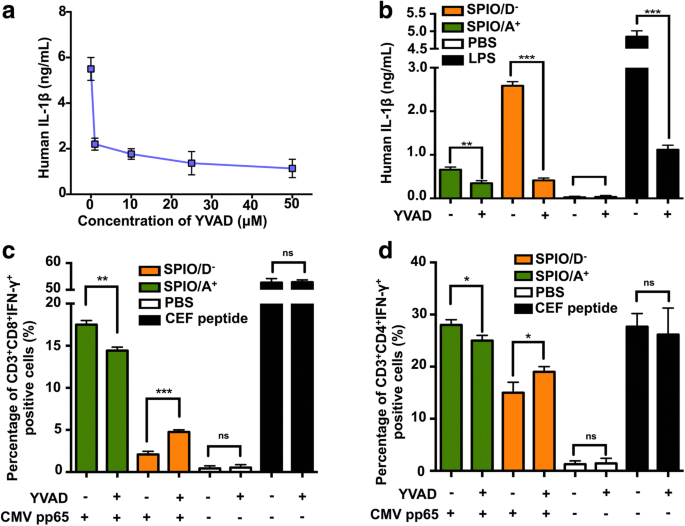
SPIO revestido com cargas opostas afeta a função das DCs humanas através da via da IL-1β. a Após o pré-tratamento por LPS por 3 h, as DCs humanas foram incubadas com concentrações crescentes de YVAD (1, 10, 25 e 50 μM) e, em seguida, os sobrenadantes foram coletados para um ELISA de IL-1β. b YVAD pode inibir a secreção de IL-1β de DCs humanas via SPIO / D - . SPIO / A + e SPIO / D - afetam a apresentação cruzada de DC influenciada por IL-1β. c CD3 + CD8 + IFN-γ + e d CD3 + CD4 + IFN-γ + As células T foram analisadas por coloração intracelular usando FCS
Discussão
A imunoterapia tem sido um foco de pesquisa em estudos clínicos e experimentais desde o desenvolvimento da biotecnologia. No entanto, os ensaios clínicos anteriores de vacinas DC tradicionais projetadas para induzir imunidade não induziram respostas imunes suficientes [26]. As nanopartículas representam um tipo de adjuvante, e podem promover a função das DCs para ativar as células T [27]. As características mais características de nosso SPIO incluem sua biocompatibilidade e sua aplicação de marcação de DCs como um adjuvante imunológico. As imagens TEM mostram que nosso SPIO tem um tamanho médio de 8,7 nm no estado seco e uma forma esférica (Fig. 1a). A coloração com azul da Prússia demonstrou que as SPIO são suscetíveis a serem fagocitadas pelas DCs (Fig. 1e), indicando a eficiência da ingestão.
Foi relatado que SPIO tem amplas aplicações biomédicas, como rotulagem celular, distribuição de drogas, ressonância magnética e hipertermia magnética [28], todas aplicações que requerem biocompatibilidade. Portanto, se as DCs marcadas com SPIO têm uma influência na apoptose das DCs deve ser investigado. Nossos dados sugeriram que não houve diferença significativa na apoptose das DCs quando as DCs foram marcadas com menos de 50 μg / mL de SPIO (Fig. 2b). No estudo a seguir, escolhemos a proteína OVA como proteína modelo e B 3 Células Z T como células T eficazes para observar se SPIO pode afetar a apresentação cruzada de DCs. Nossos resultados indicam que as DCs marcadas com SPIO podem facilitar marcadamente a apresentação cruzada de OVA e a ativação de B 3 Células Z T. As moléculas de superfície CD80 e CD86, que são marcadores de DCs maduras, são dois fatores coestimulatórios críticos necessários para a ativação de células T. A 25 μg / mL, o B 3 ativado As células Z T aumentaram com a dose de nanopartículas e eventualmente se estabilizaram (Fig. 2a). Além disso, a expressão de CD80 e CD86 na superfície das DCs atingiu um máximo (Fig. 2c). Portanto, adotamos a concentração de 25 μg / mL e verificamos que a apoptose das DCs é independente do tempo (Fig. 2d), o que comprovou as excelentes funções nanoadjuvantes e biocompatibilidade da SPIO.
Para examinar a influência de SPIO na migração de DC, usamos EGFP-DCs, que apresentou fluorescência verde sob a microscopia de fluorescência confocal, para cocultivar com 25 μg / mL SPIO por 12 h. Observamos que a expressão da fluorescência de EGFP não enfraqueceu após a marcação com SPIO. Em última análise, a migração de DCs para os órgãos linfóides secundários é o parâmetro chave para avaliar a eficácia das vacinas baseadas em DCs. Em nosso estudo anterior, nas primeiras 24 horas, as DCs marcadas com SPIO não foram detectadas em uma quantidade significativa nos linfonodos secundários [29]. Para determinar ainda mais se este fenômeno mudou com o tempo, EGFP-DCs marcados e não marcados foram coletados e injetados nas almofadas das patas de camundongos para avaliar o potencial de SPIO na migração de DCs. Nossos resultados mostraram que a fluorescência verde apareceu no ILN nos dias 4 e 7 (Fig. 3d-f), demonstrando assim que SPIO pode facilitar a migração de EGFP-DCs para os linfonodos secundários. Esta propriedade pode ser aplicada para restringir a metástase de tumores e ativar uma maior quantidade de células CTL em mais linfonodos.
Em nosso estudo, exploramos as propriedades adjuvantes de SPIO de carga oposta e o possível mecanismo subjacente às mudanças na função das DCs. A carga superficial das nanopartículas de óxido de ferro mostrou ter influência na eficiência de absorção celular [30, 31]. Em nossa pesquisa anterior, relatamos que SPIO catiônico poderia aumentar a apresentação cruzada do antígeno e, consequentemente, a ativação de células T, enquanto SPIO aniônico estava associado à autofagia. [16] Várias nanopartículas de metal podem induzir respostas inflamatórias [32, 33]. Foi relatado que nanopartículas de óxido de ferro ativam o NLRP3, rompem as membranas dos lisossomas, liberam catepsina B e induzem a secreção de IL-1β [34]. Assim, formulamos a hipótese de que as nanopartículas de SPIO com carga oposta poderiam servir a diferentes funções na estimulação da produção de IL-1β em DCs murinas e humanas. Na Fig. 4a, nossa análise de potencial zeta mostrou que SPIO revestido com APTS carregava cargas positivas, enquanto SPIO revestido com DMSA carregava cargas negativas. Sob o TEM, descobrimos que SPIO com carga diferente foram agrupados em diferentes posições no citoplasma (Fig. 4b). Para investigar a diferença entre a apresentação cruzada de SPIO / A + e SPIO / D - , exploramos a secreção de IL-1β em DCs murinas induzida por nanopartículas com cargas diferentes. Avaliamos as respostas das DCs murinas após o tratamento com SPIO / A + e SPIO / D - . Exposição a SPIO / D - induziu a ativação evidente da secreção de IL-1β em comparação com a exposição a SPIO / A + (Fig. 4c). Este fenômeno revelou parcialmente que a apresentação cruzada do antígeno induzida por SPIO / D - não é tão eficiente quanto aquele induzido por SPIO / A + . Além da análise de CPRG de aumento de B 3 Ativação de células Z T após a cocultura de DCs murinas com SPIO / A + + OVA demonstrou que nanopartículas carregadas positivamente podem ter melhor desempenho quando usadas como adjuvantes imunológicos. Receptores Toll-like (TLRs) são importantes para desencadear respostas imunes, como TLR3, e podem resultar na produção de inflamassomas, ativação da proteína caspase-1 e secreção de citocina IL-1β [35]. Para demonstrar se o TLR3 está relacionado à apresentação cruzada do antígeno influenciada pelo nosso nanoadjuvante, as DCs nocaute do TLR3 foram empregadas em nosso estudo. TLR3 - / - DCs carregados com SPIO / A + + OVA e SPIO + OVA estimularam efetivamente o B 3 Ativação de células Z T (Fig. 4d), o que indica que a molécula TLR3 em DCs foi necessária para apresentação cruzada. Coletivamente, esses resultados elucidaram que polímeros com cargas diferentes na superfície das nanopartículas devem ser considerados na investigação de seus efeitos sinérgicos nas respostas imunes ativadas.
Nossos dados mostram que as nanopartículas com carga oposta podem induzir a produção de IL-1β, enquanto SPIO / D - poderia hiperativar a secreção de IL-1β. No entanto, em comparação com SPIO / A + , SPIO / D - induziu um nível inferior de B 3 Ativação de células Z T. Portanto, especulamos que a produção aberrante de IL-1β pode contribuir para a disfunção celular em DCs. A maioria dos estudos sobre adjuvantes de nanomateriais explora DCs de camundongos, enquanto poucos usaram DCs de humanos [36, 37]. Para verificar melhor nossa hipótese, usamos YVAD, que se provou ser um inibidor eficaz da caspase-1 (Fig. 5a, b), para suprimir a secreção de IL-1β das DCs humanas. A proteína CMV pp65 foi selecionada como o antígeno modelo, e células T específicas de CEF expandidas in vitro foram usadas como células respondedoras. A proteína CMV pp65 é uma proteína imunológica dominante que prontamente ativa CD8 + e CD4 + Células T para produzir citocinas, particularmente IFN-γ [38]. O peptídeo CEF foi usado como um controle positivo para verificar se a adição de YVAD não afetaria a ativação das células T porque pode ser apresentado às células T diretamente. Curiosamente, com o uso de YVAD, as respostas das células T no SPIO / A + grupo foram ligeiramente contidos, enquanto as respostas no SPIO / D - grupo aumentou (Fig. 5c, d). A influência de IL-1β na apresentação cruzada de DC forneceu evidências conclusivas de que um baixo nível de IL-1β induzido por SPIO / A + é necessário para a apresentação cruzada do antígeno, e um alto nível de IL-1β no citosol irá inibir a função das DCs.
Conclusões
Conforme mostrado no resumo gráfico na Fig. 6, SPIO pode promover a maturação, migração e apresentação cruzada de CDs. A atividade moderada de IL-1β está parcialmente relacionada à apresentação cruzada de antígenos de DCs. Além disso, nanopartículas carregadas negativamente podem ativar o excesso de IL-1β e, subsequentemente, inibir as funções das DCs. Em resumo, nossos resultados indicam que SPIO exibe muitas propriedades biológicas e tem potencial adjuvante promissor. Essas descobertas ajudarão a identificar a escolha ideal de nanoadjuvantes para o desenvolvimento de vacinas DC no futuro.
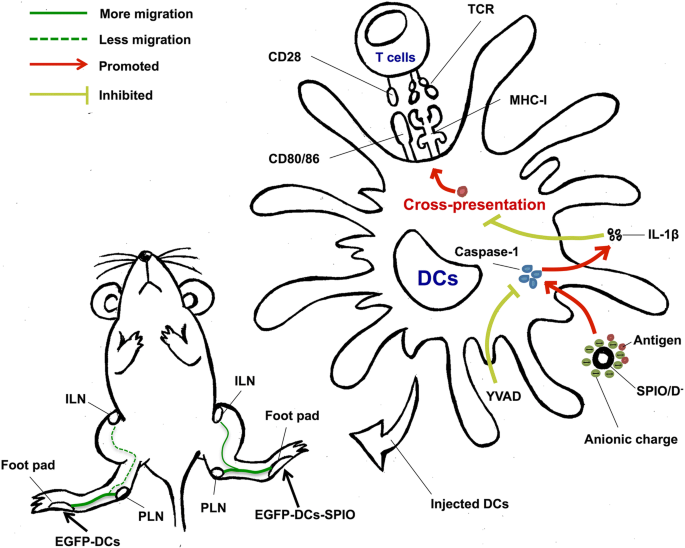
Resumo gráfico de SPIO como um nano-adjuvante para DCs. SPIO aprimora a função dos CDs, promovendo o carregamento dos CDs; assim, as DCs marcadas com SPIO podem migrar para os ILNs ativos em um órgão imune e podem oferecer uma nova abordagem na imunoterapia do câncer. SPIOs carregados com aniônicos ativam respostas protetoras de IL-1β desencadeando caspase-1 em DCs, prejudicando assim a apresentação de antígenos para células T ativas
Abreviações
- APCs:
-
Células apresentadoras de antígeno
- APTS:
-
3-aminopropiltrimetoxissilano
- ATP:
-
Trifosfato de adenosina
- CPRG:
-
Clorofenol vermelho-β-d-galactopiranosídeo
- CTL:
-
Célula T citotóxica
- DAMPs:
-
Padrões moleculares associados a danos
- DC:
-
Células dendríticas
- DMSA:
-
Ácido meso-2, 3-dimercaptosuccínico
- EGFP:
-
Proteína fluorescente verde aprimorada
- IL-1β:
-
Interleucina-1β
- MHC-I:
-
Complexo principal de histocompatibilidade classe I
- NLRP3:
-
Proteína 3 contendo domínios NACHT, LRR e PYD
- OVA:
-
Ovalbumina
- PAMPs:
-
Padrões moleculares associados a patógenos
- SPIO:
-
Nanopartículas de óxido de ferro superparamagnéticas
- SPIO / A + :
-
SPIO revestido com APTS
- SPIO / D - :
-
SPIO revestido com DMSA
- TLRs:
-
Receptores semelhantes a portagens
Catalisadores à base de platina em vários suportes de carbono e polímeros condutores para aplicações em células de combustível de metanol direto:uma revisão
Ajuste de diatomita decorada com nanoestruturas férricas hierárquicas para supercondensadores
Nanomateriais
- Integrais Definidos e o Teorema Fundamental do Cálculo
- Como a IoT remodela a indústria 4.0 e os efeitos da IoT nas PMEs
- Os efeitos da relação Li / Nb na preparação e desempenho fotocatalítico de compostos Li-Nb-O
- Influência da Água na Estrutura e Propriedades Dielétricas da Microcristalina e Nano-Celulose
- Efeitos da Espessura de Bicamada nas Propriedades Morfológicas, Óticas e Elétricas de Nanolaminados de Al2O3 / ZnO
- Os efeitos de acoplamento de polaritons de plasma de superfície e ressonâncias de dipolo magnético em metamateriais
- Fabricação e caracterização de Nano-Clips de ZnO pelo processo mediado por poliol
- Efeitos de tamanho e agregação / aglomeração de nanopartículas nas propriedades interfaciais / interfase e resistência à tração de nanocompósitos de polímero
- Diga adeus aos pontos e traços para aprimorar a mídia de armazenamento óptico
- Os efeitos de queima de plasma e oxi/combustível