


Impacto das nanopartículas de ouro no metabolismo da testosterona em microssomos hepáticos humanos
Resumo
Os complexos de nanopartícula de ouro (AuNP) -proteína corona podem alterar o metabolismo da testosterona mediada pelo citocromo P450 (CYP) (TST) ao alterar suas propriedades físico-químicas. Nós investigamos o impacto do tamanho de NP, química de superfície e corona de proteína no metabolismo de TST em microssomas de fígado humano agrupados (pHLM) empregando AuNP 40 e 80 nm funcionalizado com polietilenimina ramificada (BPEI), ácido lipóico (LA) e polietilenoglicol (PEG ), bem como corona de proteína plasmática humana (PC). A variação individual no metabolismo de TST mediado por AuNP também foi caracterizada entre HLM de doador único que continha diferentes níveis de atividades de CYP. Os efeitos inibitórios do AuNP de 40 nm e em menor grau de AuNP de 80 nm ocorreram para a produção de um total de cinco metabólitos hidroxilados de TST em pHLM, mas o PC os aliviou. Enquanto isso, o AuNP nu aumentou a produção de androstenediona. A variação interindividual no metabolismo do TST ocorreu no HLM de um único dador. Na maioria dos casos, 40 e 80 nm nu e PC AuNP suprimiram essencialmente o metabolismo de TST em concentração não inibitória, mas PC PEG-AuNP aumentou a androstenediona. Esses estudos contribuem para uma melhor compreensão do papel do AuNP como disruptor TST ao alterar o metabolismo do TST e podem ser utilizados para rastrear outros NP como um potencial disruptor endócrino.
Introdução
Nanopartículas de ouro (AuNP) têm sido amplamente empregadas na distribuição de drogas, diagnóstico médico e teranósticos de câncer, bem como em produtos de consumo, ou seja, cosméticos, embalagens de alimentos, devido às suas propriedades ópticas e físicas únicas [1,2,3]. Após a exposição a uma mistura de proteínas, os NP associam-se a proteínas e formam a corona protéica, que altera a química da superfície, as conformações de uma proteína adsorvida e as respostas biológicas subsequentes, ou seja, toxicidade de NP, absorção celular de NP, atividade catalítica do citocromo P450 (CYP ) enzimas para drogas [4,5,6,7]. Estudos in vitro com células epiteliais primárias e linhas de células cancerosas sugerem que AuNP era tóxico para hepatócitos humanos, linha celular de hepatoma C3A e células de esperma [6,7,8]. Mas a formação de proteína corona em torno de NP intensamente atenuou ou potencializou a toxicidade de AuNP de uma maneira dependente da química de superfície [6, 7]. A proteína corona interferiu na captação celular de AuNP em hepatócitos humanos, células do túbulo proximal renal, células HepG2, linha celular C3A, independentemente de seus tamanhos e cargas de superfície [6, 7, 9,10,11,12].
As enzimas CYP hepáticas estão principalmente envolvidas na síntese e / ou metabolismo de compostos endógenos e exógenos, mas uma ampla gama de agentes, ou seja, drogas, pesticidas ou NP, afetam reversamente a síntese de hormônios esteróides, metabolismo e / ou desintoxicação, resultando em efeitos farmacológicos e a função fisiológica [13,14,15,16,17]. A testosterona (TST) é um andrógeno importante e substrato específico do CYP3A4 (uma conversão principal para 6β-OH TST) de maneira regional e estereosseletiva [18]. Durante o metabolismo da fase I, o TST também é hidroxilado em 2β-OH TST pelo CYP3A4 e desalquilado em androstenediona (AD) pelo CYP2D6 [17, 19]. Estudos in vitro com hepatócitos humanos, linha celular C3A, microssomas de fígado humano (HLM) e enzimas CYP recombinantes sugeriram que AuNP nua e revestida por corona de proteína modulou uma ampla gama de enzimas CYP que incluíram CYP1A2, 2C9, 2C19, 2D6, 2E1, e 3A4 [6, 7, 20, 21]. Outro NP metálico, AgNP nu, também suprimiu a produção mediada por CYP3A4 de 6β-OH TST em HLM [22]. AuNP funcionalizado com polietilenimina ramificada (BPEI) e ácido lipóico (LA) diminuiu a atividade do CYP3A4 na linha de células C3A, mas a proteína corona plasmática humana (PC) a atenuou [7]. Em contraste, nu (sem PC) e PC BPEI-AuNP foram inibitórios para CYP2C9 e 3A4 em hepatócitos humanos, independentemente do tamanho de NP [6].
Estudos in vivo relataram que o tamanho pequeno do AuNP (4 e 13 nm) foi acumulado principalmente no fígado e no baço em camundongos BALB / c machos e induziu a expressão dos genes hepáticos Cyp1a1 e 2b [23]. Outro NP metálico, NP de óxido de zinco foram inibidores para a atividade hepática de CYP1A2, 2C11 e 3A2 em ratos machos Sprague Dawley com um aumento nas alterações patológicas no fígado [24].
Até o momento, pouco se sabe como AuNP associa o metabolismo de TST mediado por CYP (hidroxilação e desalquilação de TST) na ausência e / ou presença de corona de proteína biologicamente relevante. Os objetivos deste estudo são investigar o impacto do PC nas propriedades físico-químicas do BPEI AuNP 40 e 80 nm catiônico, LA AuNP aniônico e polietilenoglicol neutro (PEG) AuNP. O impacto do AuNP no metabolismo de TST mediado por CYP com e sem PC será caracterizado usando pHLM. A variação individual no metabolismo do TST também será estudada no HLM de um único dador que continha vários graus de enzimas CYP.
Métodos / Experimental
Químicos
O 2,3,4- 13 C 3 testosterona (CAS # 327048-83-9) e 6β-hidroxitestosterona (6β-OH TST, CAS # 62-99-7) foram obtidos de MilliporeSigma (St. Louis, MO). Testosterona (TST, CAS # 58-22-0), 2α-hidroxitestosterona (2α-OH TST, CAS # 004075-14-3), 2β-hidroxitestosterona (2β-OH TST, CAS # 10390-14-4), 6α -hidroxitestosterona (6α-OH TST, CAS # 2944-87-8), 11β-hidroxitestosterona (11β-OH TST, CAS # 1816-85-9), 15β-hidroxitestosterona (15β-OH TST, CAS # 39605-73- 7), 16α-hidroxitestosterona (16α-OH TST CAS # 63-01-4), 16β-hidroxitestosterona (16β-OH TST, CAS # 17528-90-4), 11-cetotestosterona (CAS # 564-35-2) , androstenediona (AD, CAS # 63-05-8), 4-hidroxi androstenediona (CAS # 566-48-3) e 11β-hidroxi androstenediona (CAS # 382-44-5) foram adquiridos de Steraloids (Newport, RI ) O acetonitrila e o ácido fórmico de grau LC-MS foram obtidos da Fisher Scientific (Fair Lawn, NJ), enquanto a água ultrapura foi produzida internamente pelo sistema Synergy® UV-R da Merck KGaA (Darmstadt, Alemanha). Se não for especificado, todos os outros reagentes foram adquiridos na MilliporeSigma (St. Louis, MO).
Microssomos de fígado humano
Microssomas de fígado humano agrupados (pHLM) (200 dadores, 100 homens e 100 mulheres) e um único dador de microssomas de fígado foram obtidos de Corning Inc. (Charlotte, NC). Os pHLM são agrupados pelo fornecedor, mas não por um único HLM doador. As características e a atividade enzimática do citocromo P450 (CYP) selecionada do HLM de dador único usado neste estudo são apresentadas no arquivo adicional 1:Tabela S1.
Síntese de nanopartículas de ouro
Biopure ™ 40 e AuNP esférico de 80 nm funcionalizado com polietilenimina ramificada catiônica (BPEI), ácido lipóico aniônico (LA) e polietilenoglicol neutro (PEG) foram adquiridos da nanoComposix (San Diego, CA). Os materiais do núcleo foram sintetizados através da redução do hidrato de tetracloroaurato (III) de hidrogênio (HAuCl 4 3H 2 O) em solução aquosa de carbonato de potássio e submetido ao processo de envelhecimento e filtração em fluxo tangencial (TFF). A superfície AuNP foi funcionalizada com LA ou PEG pela adição de ácido di-hidrolipóico (0,2:1, w / w ) ou PEG terminado com tiol-metoxi (Laysan Bio Inc., Arab, AL) (0,5:1, w / w ), respectivamente, com lavagem TFF e filtração estéril. Superfícies funcionalizadas com BPEI foram sintetizadas via química EDC / NHS ligando o ácido carboxílico de LA a aminas de BPEI. O BPEI não ligado foi removido com lavagem TFF e uma centrifugação subsequente.
Preparação de proteína corona de plasma humano
Plasma humano agrupado (HP, n =5) foi obtido na Biological Speciality Corp. (Colmar, PA). O AuNP de 40 e 80 nm foi incubado com plasma humano em volume de plasma fisiológico em volume de sangue total, 55% ( v / v ) em uma incubadora orbital com agitação / rotação a 37 ° C a 250 rpm por 1 h. No final da incubação, a corona de proteína plasmática humana (PC) em torno de NP foi coletada por centrifugação a 20.000 × g a 20 ° C por 20 min seguido por três lavagens com solução salina tamponada com fosfato (PBS). As proteínas não ligadas e fracamente ligadas foram descartadas por centrifugação. Os PC AuNP resultantes foram dispersos em PBS e usados para caracterização de propriedades físico-químicas e sua interação com TST.
Caracterização física de AuNP
Tamanho de partícula e propriedades de superfície foram medidos por espalhamento dinâmico de luz (DLS) e microscopia eletrônica de transmissão (TEM). Diâmetros hidrodinâmicos (D H ), e o potencial zeta de 40 e 80 nm nu (sem PC) BPEI-, LA- e PEG-AuNP em água desionizada (DI) e PC AuNP em PBS foram medidos com o Zetasizer Nano-Zs (Malvern Instruments, Worcestershire, UK) a 0 h a 25 ° C. O D H , índice de polidispersidade (PDI) e potencial zeta também foram obtidos para AuNP nu e PC em um tampão de incubação microssomal (pH 7,4) a 0 min e 45 min a 37 ° C. As amostras foram medidas cinco vezes com 11 subexecuções de 10 s cada. TEM caracterizou a morfologia de AuNP nu e PC. Todos os AuNP foram colocados em grades de cobre revestidas com formvar e visualizados em um Tecnai G2 Spirit BioTWIN com um detector Oxford (FEI Company, Hillsboro, OR) a uma tensão de aceleração de 120 kV. O conjunto de microscopia GATAN (GATAN Inc., Pleasanton, CA) mediu os diâmetros AuNP. Um espectro de absorção óptica foi medido com o leitor de microplacas multimodo Spectra Max i3 (Molecular Devices, Sunnyvale, CA).
Metabolismo in vitro da testosterona na ausência e presença de AuNP nu e PC
Estudos preliminares foram conduzidos para determinar o tempo de incubação e as concentrações de proteína microssomal para fornecer uma taxa metabólica linear para TST (uma concentração final de 10 μM). A produção de metabólitos TST foi linear de 1,3 a 9,3 mg de proteína microssomal mL -1 por até 60 min. Os ensaios metabólicos foram realizados conforme descrito na íntegra [25]. Resumidamente, pHLM em um tampão de incubação microssomal foi tratado com 10 μM TST e, subsequentemente, 40 e 80 nm nu (sem PC) AuNP foram adicionados a 0, 7, 32, 63, 143, 250, 400 e 571 μg mL -1 ; para PC AuNP pHLM 0, 7, 32, 63 e 143 μg mL −1 . Um tampão de incubação microssomal continha tampão fosfato 100 mM, MgCl 3,3 mM 2 e EDTA 1 mM (pH 7,4). A reação metabólica foi iniciada com e sem sistema de regeneração NADPH que continha NADP 0,25 mM, glicose-6-fosfato 2,5 mM e 2 U mL −1 glicose-6-fosfato desidrogenase. Após incubação de 45 min a 37 ° C, a reação foi interrompida pela adição de 4% ( v / v ) solução aquosa de ácido fosfórico (1:1, v / v ) Após uma centrifugação a 3500 rpm por 20 min, um sobrenadante de amostra foi coletado e armazenado a -20 ° C até o uso posterior. Além disso, um único doador HLM em um tampão de incubação microssomal foi tratado com 10 μM de TST seguido de incubação com 63 μg mL −1 de todos nus e PC AuNP por 45 min a 37 ° C. No final da incubação, a amostra foi processada e armazenada a -20 ° C como acima.
Padrões e preparação de amostra
Soluções de estoque padrão primárias de TST, seus metabólitos e 13 C 3 TST marcado como um padrão interno (ISTD) foram preparados em metanol a uma concentração de 1 mM e armazenados a -20 ° C até uso posterior. As concentrações para soluções padrão de trabalho de TST e seus metabólitos foram 0,01, 0,05, 0,1, 0,5, 1, 5, 10, 50, 100 e 200 μM com diluições em série da solução estoque primária. Para calibradores padrão, uma alíquota de 50 μL de cada solução padrão de trabalho foi adicionada a 450 μL de tampão de reação, resultando em diluições de 1:10, enquanto 0,1 μM de solução ISTD também foi preparada usando solução aquosa de ácido fosfórico a 4%. Amostras de controle de qualidade (QC) foram preparadas em concentrações de 0,01, 0,05 e 0,1 μM.
Após o descongelamento, as amostras foram centrifugadas a 3500 rpm por 20 min em temperatura ambiente. O sobrenadante foi enriquecido com 50 μL de 0,1 μM de ISTD e submetido a placa de eluição de 96 poços Oasis PRIME HLB e uma placa de coleta no processador Waters de pressão positiva de 96 a 80 psi por 1–2 min. (Waters Corp., Milford, MA). Após lavagem com 300 μL de água com 5% de metanol e eluição com 50 μL de mistura de acetonitrila / metanol (90/10, v / v ), o eluente resultante foi diluído em 50 μL de água (um volume final de 100 μL) e sujeito a cromatografia líquida-espectrometria de massa (LC-MS / MS).
Cromatografia líquida-espectrometria de massa
Todas as amostras foram separadas em coluna Waters UPLC HSS T3 (2,1 × 50 mm, 1,8 μm) com sistema de cromatografia líquida Waters Acquity Ultra Performance com detector triplo quadrupolo (UPLC TQD) (Waters Corp., Milford, MA). As fases móveis A e B foram 0,1% de ácido fórmico em água e 0,1% de ácido fórmico em metanol, respectivamente. Um método de gradiente LC foi usado a uma taxa de fluxo de 600 μL min −1 por 0–8,4 min. O gradiente foi de 0-1 min (30% B), 1-3 min (para 50% B), 3-3,5 min (50% B), 3,5-7 min (para 80% B), 7-7,01 min ( a 98% B), 7,01-7,5 min (98% B) e 7,51-8,4 min (30% B). As condições de MS foram descritas resumidamente a seguir. A fonte de ionização foi operada em eletrospray positivo (ESI + ) modo com tensão capilar 4000 V; para temperatura da fonte 150 ° C; e para temperaturas de dessolvatação 450 ° C. As taxas de fluxo do gás de dessolvatação (N 2 ), gás de cone (N 2 ), e o gás de colisão (argônio) foram 900 L h −1 , 100 L h −1 , e 0,1 mL min −1 , respectivamente. O tipo de varredura foi o monitoramento de reação múltipla (MRM) e o tempo de execução do MS foi de 8,4 min. As transições MRM usadas para a análise foram resumidas no arquivo adicional 1:Tabela S2 e Fig. 2. O volume de injeção foi de 2 μL e a coluna foi mantida a 50 ° C durante toda a análise. Todos os métodos de quantificação foram baseados em uma curva de calibração de sete pontos nas faixas de concentração de 0,001 a 20 μM. O limite de detecção (LOD) e o limite de quantificação (LOQ) de 0,001 μM e 0,005 μM foram estabelecidos para TST e metabólitos alvo.
Análise estatística
Os efeitos dos dispersantes em D H e PDI de nu e PC no AuNP foram avaliados usando o t do aluno teste com uma distribuição bicaudal. Metade da concentração inibitória máxima (IC 50 ) e metade da concentração máxima de ativação (EC 50 ) de AuNP para a produção de metabólitos de TST dependentes de CYP em pHLM foram determinados ajustando uma equação de Hill com inclinação variável aos dados observados usando GraphPad Prism®. Uma análise de variância unilateral (ANOVA) foi conduzida usando GraphPad Prism® para avaliar os efeitos do tratamento com AuNP no metabolismo do TST em HLM de doador único. Quando os efeitos foram significativos, uma comparação múltipla foi realizada com o teste de diferença significativa honesta (HSD) de Tukey com nível de significância de 5%. Coeficiente de correlação de Pearson ( r ) entre a atividade de CYP de HLM de dador único e a produção de metabólitos de TST dependentes de CYP foi determinada usando GraphPad Prism® versão 6.07 (La Jolla, CA).
Resultados e discussão
Caracterização físico-química de proteína plasmática nua e humana Corona AuNP
O impacto da corona da proteína plasmática humana (PC) no tamanho de NP, carga de superfície e morfologia, bem como uma propriedade espectral, foi caracterizado usando DLS, TEM e espectroscopia UV-Vis (Fig. 1). As imagens TEM demonstraram que todos os nus (sem PC) e PC AuNP, exceto para 40 e 80 nm PC BPEI-AuNP foram monodispersos com a distribuição de tamanho estável e faixas de espectro UV-Vis exclusivas (520-557 nm) (Fig. 1a, b) . PC distintos em torno de AuNP em PBS também foram encontrados por TEM. A agregação da polietilenimina ramificada revestida com PC de 40 e 80 nm (BPEI) -AuNP em PBS a 0 min a 25 ° C correlacionou múltiplos picos na distribuição de tamanho e redshifts dos espectros de absorção em relação ao BPEI-AuNP nu (Fig. 1b). O diâmetro hidrodinâmico (D H ) os valores de 40 e 80 nm PC BPEI-AuNP dissolvido em PBS a 0 min a 25 ° C e em tampão de incubação microssomal a 0 e 45 min a 37 ° C não foram determinados por DLS, juntamente com múltiplos picos na distribuição de tamanho. O D H os valores do BPEI- e LA-AuNP nu de 40 nm e PC PEG-AuNP e do BPEI-AuNP nu de 80 nm em tampão de incubação microssomal aumentaram substancialmente até 45 min a 37 ° C, enquanto seu valor diminuiu para o PC LA de 80 nm -AuNP (Tabela 1). O índice de polidispersidade (PDI) de 40 nm nu e PC PEG-AuNP e 80 nm PC PEG-AuNP aumentou em 45 min a 37 ° C. Além disso, os valores de potencial zeta (z) do BPEI-AuNP de 40 e 80 nm nu e de PEG-AuNP de 40 nm nu diminuíram substancialmente ao longo do tempo. Estudo anterior relatou que AuNP (7 e 70 nm) associado com proteínas microssômicas do fígado humano alterou a absorbância máxima característica na faixa de UV-visível [21]. Esses resultados foram apoiados por estudos recentes em nosso laboratório de que PC e corona de albumina sérica humana alteraram o tamanho de NP, desvio para o vermelho da absorbância máxima e morfologia, independentemente do meio de dissolução e do tempo de incubação [6, 7, 10, 26]. Com base nas alterações nas propriedades físico-químicas de NP mediadas por PC e na função enzimática das atividades de CYP [6, 7], o efeito potencial do AuNP de 40 e 80 nm no metabolismo de TST microssomal hepático humano mediado por CYP foi investigado na presença de um método mais biológico PC relevante.

Micrografias eletrônicas de transmissão de ( a ) o AuNP nu de 40 e 80 nm em água deionizada e b o 40 e 80 nm PC AuNP em PBS a 0 h a 25 ° C, espectro de absorção de UV (inserção superior) e a distribuição de espalhamento de luz dinâmica (inserção inferior). As setas indicam a formação do PC. BPEI polietilenimina ramificada, LA ácido lipóico, PEG polietilenoglicol, ND não determinado, PC corona de proteína plasmática humana, nua sem PC
Metabolismo de testosterona mediado por AuNP em microssomos hepáticos humanos agrupados
Um total de 11 metabólitos de TST foram selecionados e seis metabólitos foram encontrados em microssomas de fígado humano agrupados (pHLM) a 10 μM de TST. A análise LC-MS / MS direcionada de TST e os seis metabólitos selecionados é demonstrada na Fig. 2. A lista dos metabólitos selecionados incluiu cinco metabólitos TST hidroxilados (2β-OH TST, 6β-OH TST, 15β-OH TST, 16α -OH TST, 16β-OH TST) e um metabólito desalquilado (androstenediona, AD). Isso se correlaciona com os estudos anteriores usando hepatócitos humanos e HLM que TST foi principalmente hidroxilado em 6β-OH TST e em menor grau 2β-OH TST, 15β-OH TST, 16α-OH TST e 16β-OH TST, bem como um metabólito desalquilado, AD [17, 19, 27].
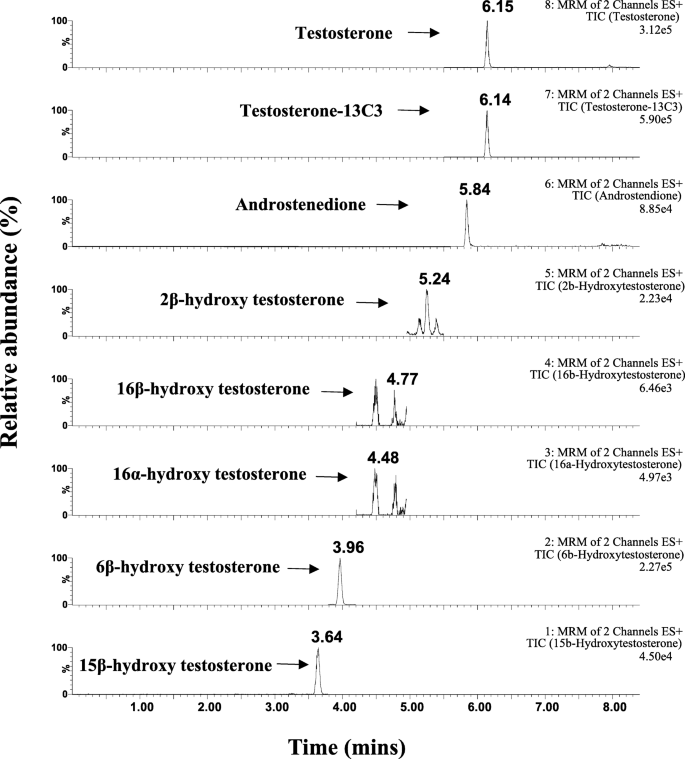
O cromatograma de íons extraídos (XIC) para testosterona (TST), 13 TST marcado com C3, androstenediona, 2β-hidroxi testosterona (2β-OH TST), 16α-hidroxitestosterona (16α-OH TST), 16β-hidroxitestosterona (16β-OH TST), 6β-hidroxitestosterona (6β-OH TST) e 15β -hidroxitestosterona (15β-OH TST) produzida em HLM agrupado na concentração final de 10 μM de testosterona na presença de NADPH por 45 min a 37 ° C. HLM microssomas de fígado humano, NADPH um fosfato de dinucleotídeo de nicotinamida adenina reduzido
O resultado da coincubação de TST com o AuNP nu de 40 e 80 nm em pHLM foi mostrado nas Figs. 3 e 4. Todos os AuNP nus de 40 e 80 nm alteraram a produção de 2β-OH TST, 6β-OH TST e 15β-OH TST em pHLM com grau variável de inibição (Fig. 3a-f). Metade da concentração inibitória máxima (IC 50 ) os valores de 40 nm BPEI-AuNP para a produção de 6β-OH TST foi de 416 μg mL −1 ; para 80 nm BPEI-AuNP 438 μg mL −1 ; e para 80 nm PEG-AuNP 387 μg mL −1 (Fig. 3c, d). Para produção de 15β-OH TST, IC 50 valores de 40 nm BPEI-AuNP foi 1113 μg mL −1 ; para 80 nm BPEI-AuNP 415 μg mL −1 ; e para 80 nm PEG-AuNP 480 μg mL −1 (Fig. 3e, f). Estes resultados foram apoiados por estudos in vitro com linhas de células de câncer humano e tecidos hepáticos que um NP metálico, AgNP e NP de silício poroso impediram a produção de TST de 6β-OH em células Caco2 de adenocarcinoma colorretal epitelial humano, células de carcinoma hepatocelular HepG2 e microssomas de fígado humano [22, 28]. O AuNP nu de 40 e 80 nm não foi inibidor da produção de 16α-OH TST e 16β-OH TST, exceto para o BPEI-AuNP de 40 nm que suprimiu a produção de 16β-OH TST na concentração mais alta (517 μg mL - 1 ) (Fig. 4a-d). O BPEI-AuNP de 40 e 80 nm aumentou a produção de androstenediona (AD) na concentração mais alta com as taxas metabólicas correspondentes de 4,3 e 2,1 pmol mg de proteína −1 min −1 , respectivamente em comparação com o controle (0,7 pmol mg de proteína −1 min −1 ) (Fig. 4e, f). Mas o LA-AuNP de 80 nm foi inibidor para a produção de AD com IC 50 valor de 233 μg mL −1 . Estes resultados indicaram que o AuNP nu mediou a produção do metabólito TST selecionado no revestimento de superfície e de maneiras dependentes do tamanho. Além disso, todos os 40 e 80 nm PC AuNP não foram inibitórios para a produção de seis metabólitos selecionados de TST em pHLM em até 143 μg mL −1 , independentemente dos revestimentos de superfície (Figs. 5 e 6). Especialmente, o PC aliviou a inibição mediada por BPEI-AuNP de 40 nm da produção de 6β-OH TST e 15β-OH TST em concentrações mais altas (32 μg mL −1 a 143 μg mL −1 ) (Figs. 3c, e e 5c, e). Estes resultados exibiram que BPEI-, LA- e PEG-AuNP nus de 40 e 80 nm diminuíram a hidroxilação de TST (2β-OH TST, 6β-OH TST e 15β-OH TST) de uma maneira dependente da dose (Fig. 7 ) Além disso, o BPEI-AuNP nu de 40 e 80 nm aumentou a produção de AD, mas o primeiro diminuiu 16β-OH TST. Um estudo in vitro relatou que a produção de 6β-OH TST mediada principalmente por CYP3A4 foi inibida por nanotubos de carbono de parede única (SWCNT) de uma maneira dependente da dose, mas a corona de albumina sérica bovina a aliviou [17, 29]. Nosso laboratório relatou recentemente que o BPEI-AuNP de 40 e 80 nm nu e PC serviram como um inibidor para CYP1A2, 2C9 e 3A4 nos níveis celular e transcricional [6, 7]. O estudo in vivo relatou que o PEG-AuNP (4 e 13 nm) se acumulou principalmente no fígado em camundongos BALB / c machos e níveis de transcrição alterados dos genes hepáticos Cyp1a1 e 2b [23]. Ratinhos ICR machos com i.v. injeção de PEG-NH 2 -AuNP exibiu NP aumentado o nível de TST no plasma sem a morfologia e fertilidade do esperma [30]. Outro NP metálico, dióxido de titânio (TiO 2 ) foi acumulado nos testículos de camundongos machos CD1 e diminuiu a expressão de cyp1b1 e 2e1 [31]. Estudo de epidemiologia relatou que homem adulto na clínica de infertilidade de Massachusetts mostrou níveis plasmáticos de TST baixos comparados com um alto nível de 3,5,6-tricloro-2-piridinol (TCP) derivado de alta exposição a clorpirifós (CFS), um desregulador endócrino conhecido e um inibidor do metabolismo de TST mediado por CYP [32]. Estudo anterior relatou que os inseticidas desreguladores endócrinos conhecidos, CFS, CFS oxon, fonofos, forato, dietiltoluamida (DEET) e permetrina inibiram e / ou ativaram substancialmente a produção dos metabólitos TST hidroxilados e / ou desalquilados, ou seja, 2β-OH TST, 6β-OH TST, 15β-OH TST e AD e 4-hidroxi AD em fígado humano [17]. Com isso dito, é razoável postular que AuNP pode ser um desregulador endócrino potencial ao mediar uma inibição e / ou potências de ativação contra o metabolismo de TST mediado por CYP.
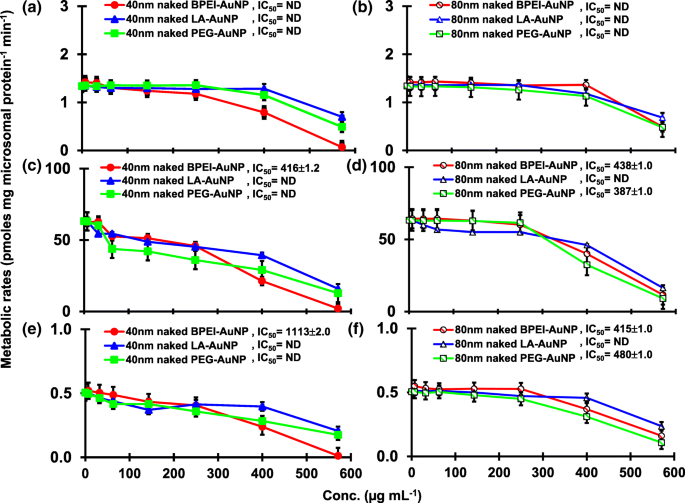
Efeito inibitório do AuNP nu (sem PC) na produção de 2β-OH TST ( a , b ), 6β-OH TST ( c , d ) e 15β-OH TST ( e , f ) em pHLM. Os dados representam a média ± D.P. ( n =3). IC 50 , metade da concentração inibitória máxima; pHLM , microssomas de fígado humano agrupados; ND , não determinado pelo ajuste de uma equação de Hill com inclinação variável aos dados observados usando GraphPad Prism®; PC , proteína plasmática humana; BPEI , polietilenimina ramificada; LA , ácido lipoico; PEG, polietilenoglicol; Conc, concentração; 2β-OH TST, 2β -hidroxitestosterona; 6β-OH TST, 6β -hidroxitestosterona; 15β-OH TST, 15β -hidroxitestosterona
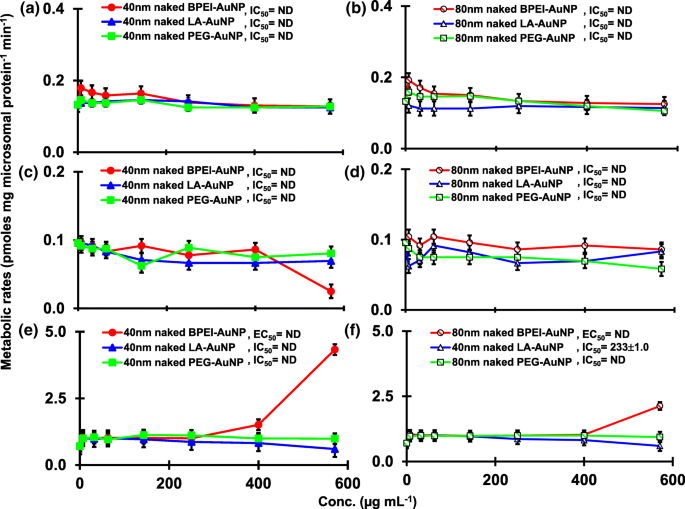
Um efeito inibitório e estimulador do AuNP nu (sem PC) na produção de 16β-OH TST ( a , b ), 16β-OH TST ( c , d ) e AD ( e , f ) em pHLM. Os dados representam a média ± D.P. ( n =3). IC 50 , metade da concentração inibitória máxima; CE 50 , metade da concentração máxima de ativação; pHLM , microssomas de fígado humano agrupados; ND , não determinado pelo ajuste de uma equação de Hill com inclinação variável aos dados observados usando GraphPad Prism®; PC , proteína plasmática humana; BPEI , polietilenimina ramificada; LA , ácido lipoico; PEG , polietileno glicol; Conc, concentração; 16α-OH TST, 16α -hidroxitestosterona; 16β-OH TST, 16β -hidroxitestosterona; AD , androstenediona.
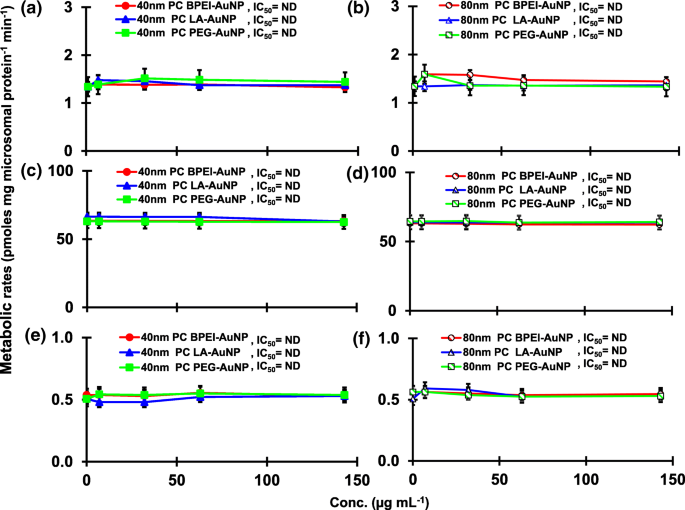
Efeitos de PC AuNP na produção de 2β-OH TST ( a , b ), 6β-OH TST ( c , d ) e 15β-OH TST ( e , f ) em pHLM. Os dados representam a média ± D.P. ( n =3). IC 50 , metade da concentração inibitória máxima; pHLM , microssomas de fígado humano agrupados; ND , não determinado pelo ajuste de uma equação de Hill com inclinação variável aos dados observados usando GraphPad Prism®; PC , corona de proteína plasmática humana; BPEI , polietilenimina ramificada; LA , ácido lipoico; PEG , polietileno glicol; Conc, concentração; 2β-OH TST, 2β -hidroxitestosterona; 6β-OH TST, 6β -hidroxitestosterona; 15β-OH TST, 15β-hidroxitestosterona.
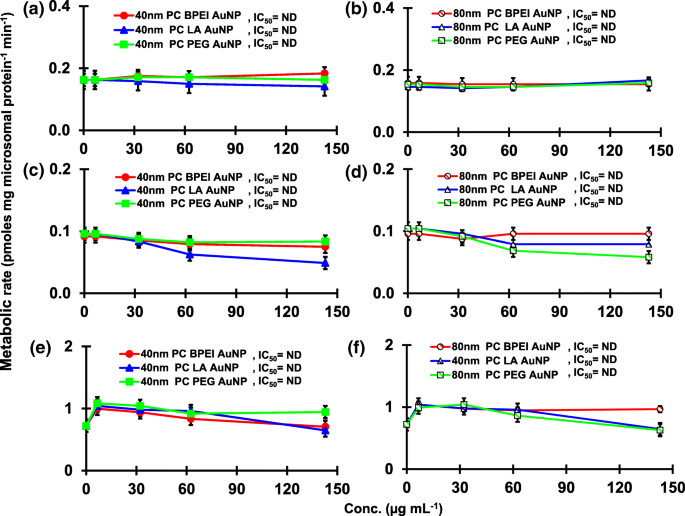
Efeitos de PC AuNP na produção de 16α-OH TST ( a , b ), 16β-OH TST ( c , d ) e AD ( e , f ) em pHLM. Os dados representam a média ± D.P. ( n =3). IC 50 , metade da concentração inibitória máxima; pHLM , microssomas de fígado humano agrupados; ND , não determinado pelo ajuste de uma equação de Hill com inclinação variável aos dados observados usando GraphPad Prism®; PC , corona de proteína plasmática humana; BPEI , polietilenimina ramificada; LA , ácido lipoico; PEG , polietileno glicol; Conc, concentração; 16α-OH TST, 16α -hidroxitestosterona; 16β-OH TST, 16β -hidroxitestosterona; AD , androstenediona.
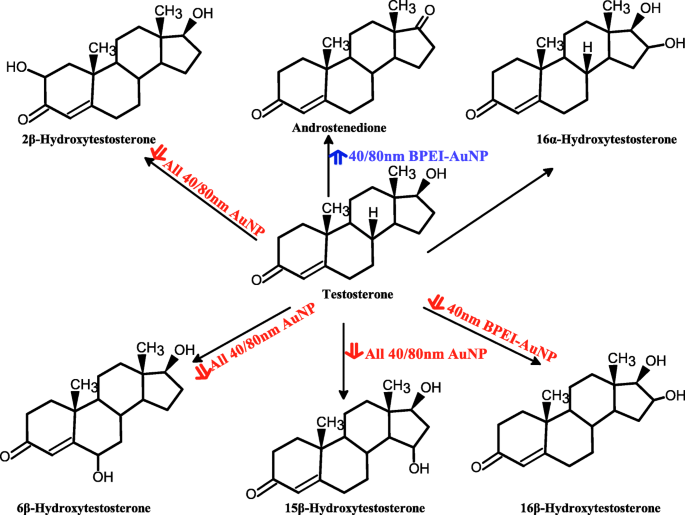
Esquema proposto de metabolismo de testosterona em microssomas hepáticos humanos combinados e produção de metabólitos mediada por AuNP. AuNP nanopartículas de ouro, BPEI polietilenimina ramificada, LA ácido lipóico, PEG polietilenoglicol, PC corona de proteína plasmática humana. Seta vermelha, efeito inibitório; seta azul, efeito estimulador
Metabolismo da TST em microssomos de fígado humano de dador único e sua modulação por AuNP
A produção de seis metabólitos selecionados foi caracterizada com um único doador HLM isolado de doadores com vários graus de atividade de CYP na concentração não inibitória baseada em pHLM de 40 e 80 nm nu e PC AuNP (10 μg mL −1 ) Uma variação individual no metabolismo TST foi observada entre três diferentes HLM de doador único (arquivo adicional 1:Figura S1). A relação entre a atividade catalítica de cada enzima CYP e a produção de metabólitos derivados de TST foi caracterizada em três diferentes HLM de doador único que continha uma baixa, uma média e uma alta atividade catalítica de CYP (Arquivo adicional 1:Tabela S1). A produção de 6β-OH TST correlacionou-se positivamente com a atividade de CYP2C19 ( r =0,99 e p =0,01) e CYP3A4 ( r =0,99 e p =0,03) dentro de indivíduos (Arquivo adicional 1:Figura S2). A produção de AD correlacionou-se negativamente com CYP4A11 ( r =- 0,98 e p =0,04) (Arquivo adicional 1:Figura S3). Esses resultados foram consistentes com um estudo anterior relatado que o CYP3A4 e o CYP2D6 desempenharam um papel fundamental na produção de um metabólito principal do TST, 6β-OH TST e AD, respectivamente [17]. Este estudo também sugeriu que uma variação individual no metabolismo dependente de CYP do TST depende dos genótipos das enzimas CYP e de suas atividades fenotípicas. Um estudo anterior relatou que polimorfismos e fenótipos de CYP são as principais características na função de CYP e resultam em fenótipos farmacogenéticos categóricos como metabolizadores fracos, intermediários, extensos e ultrarápidos, contribuindo para a suscetibilidade individual a reações adversas a medicamentos e / ou eficácia de medicamentos e sugestão de dose esse fraco metabolizador da enzima CYP, ou seja, CYP3A4, pode ser suscetível ao metabolismo de TST por exposição ao inibidor de CYP, AuNP [33].
Conforme mostrado nas Figs. 8 e 9, a coincubação de TST com AuNP em concentração não inibitória causou um aumento e / ou diminuição no metabolismo de TST mediado por CYP entre HLM de doador único em função do tamanho e modificação de mudança de superfície. ANOVA indicou que mudanças significativas pelo tamanho do AuNP ( p <0,0001), revestimentos de superfície ( p <0,0001), e formação de PC ( p <0,0001) foram observados para a produção de seis metabólitos selecionados de TST em HLM de doador único (HDA1, HDB2 e HDC3).
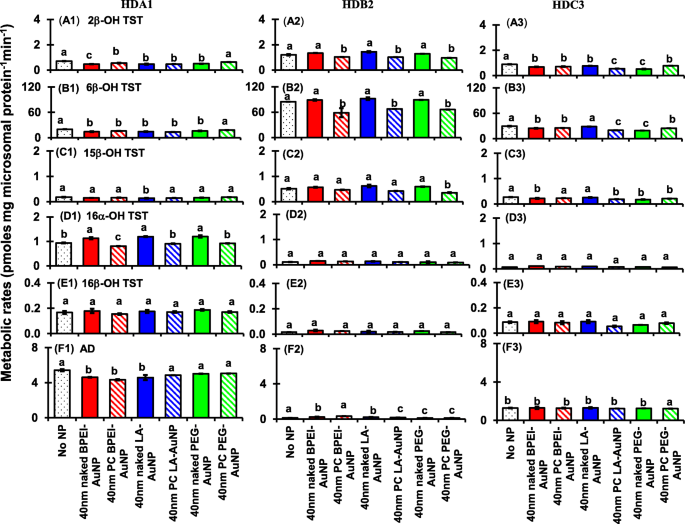
Efeitos de 40 nm nu e PC AuNP na produção de 2β-OH TST (A1 – A3), 6β-OH TST (B1 – B3), 15β-OH TST (C1 – C3), 16α-OH TST (D1 – D3 ), 16β-OH TST (E1 – E3) e AD (F1 – F3); em três diferentes HLM de doador único (HDA1, HDB2 e HDC3). Means followed by the same letter were not significantly different for each nanoparticle (Tukey’s honest significant difference =5%). Naked no PC, PC human plasma protein corona, HLM human liver microsomes, BPEI polietilenimina ramificada, LA ácido lipóico, PEG polyethylene glycol, 2β -OH TST 2β-hydroxytestosterone, 6β -OH TST 6β-hydroxytestosterone, 15β -OH TST 15β-hydroxytestosterone, 16α -OH TST 16α-hydroxytestosterone, 16β -OH TST 16β-hydroxytestosterone, AD androstenedione

Effects of 80 nm naked and PC AuNP on the production of 2β-OH TST (A1–A3), 6β-OH TST (B1–B3), 15β-OH TST (C1–C3), 16α-OH TST (D1–D3), 16β-OH TST (E1–E3), and AD (F1–F3); in three different single donor HLM (HDA1, HDB2, and HDC3). Means followed by the same letter were not significantly different for each nanoparticle (Tukey’s honest significant difference =5%). Naked no PC, PC human plasma protein corona, HLM human liver microsomes, BPEI polietilenimina ramificada, LA ácido lipóico, PEG polyethylene glycol, 2β -OH TST 2β-hydroxytestosterone, 6β -OH TST 6β-hydroxytestosterone, 15β -OH TST 15β-hydroxytestosterone, 16α -OH TST 16α-hydroxytestosterone, 16β -OH TST 16β-hydroxytestosterone, AD androstenedione
All 40 nm naked and PC AuNP decreased both 2β-OH TST and 6β-OH TST production in HDA1 and HDC3 except for PC PEG-AuNP in the former and naked LC-AuNP in the latter, whereas in HDB2, only PC AuNP potentiated inhibition of their productions, irrespective of surface coatings (Fig. 8(A1–B3)). The 40 nm naked LA-AuNP was an inhibitor for 15β-OH TST production in HDA1; for HDB2 PC PEG-AuNP; and for HDC3 naked BPEI- and PEG-AuNP and PC LA- and PC PEG-AuNP (Fig. 8(C1–C3)). All 40 nm naked AuNP were an activator for 16α-OH TST production in HDA1 but PC attenuated it except for PC BPEI-AuNP which potentiated its inhibition (Fig. 8(D1)). All 40 nm naked and PC AuNP did not influence the production of 16β-OH production within individuals (Fig. 8(E1–E3)). AD production was modulated by the 40 nm naked and PC AuNP with varying degrees of inhibition except for PC PEG-AuNP which served an activator in HDC3 (Fig. 8(F1–F3)).
The 80 nm naked and PC AuNP-mediated inhibition for 2β-OH TST production was observed within individuals except for 80 nm naked and PC PEG-AuNP which served as the activators in HDB2 and HDC3, respectively (Fig. 9(A1–A3)). These results were not consistent with all naked and PC 40 nm AuNP-mediated inhibition for 2β-OH TST, irrespective of surface coatings (Fig. 8(A1–A3)). For 6β-OH TST, the 80 nm naked AuNP were the activators except for LA-AuNP but PC potentiated its inhibition in HDB2 (Fig. 9(B2)), which was similar to the 40 nm naked and PC AuNP-mediated inhibition and/or activation, respectively (Fig. 8(B2)). For 15β-OH TST, 80 nm naked BPEI- and PEG-AuNP and PC PEG-AuNP were the activators in HDB2 and in HDC3, respectively (Fig. 9(C2 and C3)). The 80 nm naked AuNP increased 16α-OH TST production in HDA1, irrespective of surface coatings but PC attenuated it except for PC LA-AuNP which was an inhibitor (Fig. 9(D1)). This is similar to the 40 nm naked and PC AuNP-mediated activation and attenuation for 16α-OH TST production in HDA1 (Fig. 8(D1)). All 80 nm naked and PC were not inhibitory to 16β-OH TST production within all individuals, irrespective of surface coatings (Fig. 9(E1–E3)). These results were consistent with the 40 nm naked and PC-mediated its production within individuals (Fig. 8(E1–E3)). For AD production, the 80 nm naked BPEI- and PEG-AuNP were the inhibitors but PC attenuated and vice versa with naked and PC LA-AuNP in HDA1 (Fig. 9(F1)). The 80 nm naked and PC AuNP decreased its production in HDC3 except for PC PEG-AuNP, which was an activator (Fig. 9(F3)). This study strongly suggests that AuNP interaction with CYP enzymes in HLM cause a decrease and/or increase in TST conversion to hydroxylated and dealkylated metabolites within individuals and the presence of PC played the inhibitive or protective role. In vivo study reported that the male CD-1 mice orally administrated with ketoconazole, a noncompetitive CYP3A4 inhibitor showed that a decrease in serum TST level, gonadal TST secretion, and hepatic TST hydroxylation activity that included 6β-OH TST, 15α-OH TST, 15β-OH TST, and 16β-OH TST [34]. In vitro studies with human hepatocyte, C3A cell line, HepG2 cell line, HLM, and recombinant CYP enzymes suggested that AuNP modulated the activity of various CYP enzymes that included CYP1A2, 2C9, 2C19, 2D6, 2E1, and 3A4 [6, 7, 20, 21]. PC and human serum albumin corona mitigated an inhibitory effect of BPEI- and LA-AuNP on CYP1A2, 2C9, and 3A4 enzyme activity in human hepatocytes and C3A cell line [6, 7]. That being said, it may be rational to propose that AuNP interference with CYP enzymes relates individual susceptibility to unexpected toxicological effects that may result in an altered circulating TST level tied to endocrine disrupting substance and/or drug-drug interaction sharing the same CYP enzymes [35].
Conclusões
These studies exhibit that AuNP interaction with PC definitely modulate CYP-dependent metabolism of TST in HLM derived from a large donor pool that better represents the average American population. The 40 nm naked (no PC) AuNP and to a lesser degree 80 nm naked AuNP inhibited TST hydroxylation but activated TST dealkylation at high concentration. Cationic BPEI-AuNP withheld the production of 6β-OH TST and 15β-OH TST in pooled HLM but the presence of a more biologically relevant PC alleviated their adverse effects as function of size and surface charge modification. In most cases, the 40 and 80 nm naked and PC AuNP are essentially inhibitory to TST metabolism in single donor HLM in a surface chemistry-dependent manner at the noninhibitory concentration. In addition, PC PEG-AuNP caused an activation of AD production in HDC3, irrespective of size. These results may indicate that individual variations in AuNP-mediated TST metabolism could be a factor for their toxicity and could be utilized to identify vulnerable subgroup to TST-disrupting NP.
Disponibilidade de dados e materiais
All data generated or analyzed during this study are included in this article and its supplementary information file.
Abreviações
- 11β-OH TST:
-
11β-hydroxytestosterone
- 15β-OH TST1:
-
5β-hydroxytestosterone
- 16α-OH TST:
-
16α-hydroxytestosterone
- 16β-OH TST:
-
16β-hydroxytestosterone.
- 2α-OH TST:
-
2α-hydroxytestosterone
- 2β-OH TST:
-
2β-hydroxytestosterone
- 6α-OH TST:
-
6α-hydroxytestosterone
- 6β-OH TST:
-
6β-hydroxytestosterone
- AD:
-
Androstenedione
- AgNP:
-
Silver nanoparticles
- ANOVA:
-
Análise de variância unilateral
- AuNP:
-
Nanopartículas de ouro
- BPEI:
-
Polietilenimina ramificada
- CFS:
-
Chlorpyrifos
- CYP:
-
Citocromo P450
- DEET:
-
Diethyltoluamide
- DH :
-
Diâmetros hidrodinâmicos
- DI:
-
Água desionizada
- DLS:
-
Espalhamento de luz dinâmico
- EC50 :
-
Half maximal activation concentration
- EDC/NHS:
-
1-Ethyl-3-(3-dimethylaminopropyl) carbodiimide/N-hydroxysuccinimide
- ESI + :
-
Electrospray positive
- HLM:
-
Human liver microsomes
- HSD:
-
Tukey’s honest significant difference
- IC50 :
-
Half maximal inhibitory concentration
- ISTD:
-
Internal standard
- LA:
-
Lipoic acid
- LC-MS/MS:
-
Liquid chromatography-mass spectrometry
- LOD:
-
Limite de detecção
- LOQ:
-
Limit of quantitation
- MRM:
-
Multiple reaction monitoring
- NADP:
-
Nicotinamide adenine dinucleotide phosphate
- NADPH:
-
Reduced NADP
- naked:
-
No PC
- NP:
-
Nanopartículas
- PBS:
-
Salina tamponada com fosfato
- PC:
-
Corona de proteína plasmática humana
- PDI:
-
Índice de polidispersidade
- PEG:
-
Polietileno glicol
- pHLM:
-
Pooled human liver microsomes
- QC:
-
Quality control
- SWCNT:
-
Nanotubo de carbono de parede simples
- TEM:
-
Microscopia eletrônica de transmissão
- TFF:
-
Filtragem de fluxo tangencial
- TiO 2 :
-
Dióxido de titânio
- TST:
-
Testosterona
- UPLC TQD:
-
Ultra performance liquid chromatography system with Triple quadrupole Detector
Influência dominante da dispersão da rugosidade da interface no desempenho de lasers em cascata quântica GaN Terahertz
Papel fotocatalítico flexível com Cu2O e Nanopartículas de ZnO decoradas com nanopartículas de Ag para fotodegradação de corante orgânico à luz visível
Nanomateriais
- Nanopartículas de semicondutor
- Nanopartículas plasmônicas
- Nanopartículas de ouro para sensores quimio
- Nanopartículas multifuncionais de ouro para aplicações diagnósticas e terapêuticas aprimoradas:uma revisão
- Nanobiossensor de ouro com base na ressonância de plasma de superfície localizada é capaz de diagnosticar a brucelose humana, apresentando um método rápido e acessível
- Toxicidade potencial para fígado, cérebro e embrião de nanopartículas de dióxido de titânio em camundongos
- Estabilidade aprimorada de nanopartículas magnéticas de ouro com poli (ácido 4-estirenossulfônico-ácido co-maleico):propriedades ópticas sob medida para detecção de proteínas
- Poliglicerol hiper-ramificado modificado como dispersante para controle de tamanho e estabilização de nanopartículas de ouro em hidrocarbonetos
- Promoção do crescimento celular SH-SY5Y por nanopartículas de ouro modificadas com 6-mercaptopurina e um peptídeo penetrador de neurônio
- Tatuagem feita de nanopartículas de ouro revoluciona o diagnóstico médico