


Biocompatibilidade de nanocarreadores de lipossomas na orelha interna de rato após administração intratimpânica
Resumo
Os nanocarreadores de lipossomas (LPNs) são potencialmente o futuro da terapia do ouvido interno devido à sua alta capacidade de carga de drogas e captação eficiente no ouvido interno após uma administração intratimpânica minimamente invasiva. No entanto, faltam informações sobre a biocompatibilidade dos LPNs no ouvido interno. O objetivo do presente estudo é documentar a biocompatibilidade dos LPNs na orelha interna após o parto intratimpânico. LPNs com ou sem ácido gadolínio-tetra-azaciclo-dodecano-tetra-acético (Gd-DOTA) foram entregues aos ratos por meio de injeção transtimpânica. A distribuição dos LPNs contendo Gd-DOTA no ouvido médio e interno foi rastreada in vivo por meio de ressonância magnética. A função das barreiras da orelha média e interna foi avaliada por meio de ressonância magnética com gadolínio. A função auditiva foi medida por meio da resposta auditiva de tronco encefálico (ABR). A potencial resposta inflamatória foi investigada pela análise da secreção de glicosaminoglicanos e ácido hialurônico e da expressão de CD44 e TLR2 na orelha interna. A apoptose potencial foi analisada usando transferase terminal (TdT) para marcar as quebras 3′OH livres nas fitas de DNA de células apoptóticas com TMR-dUTP (coloração TUNEL). Como resultado, LPNs entraram na orelha interna de forma eficiente após a injeção transtimpânica. A injeção transtimpânica de LPNs com ou sem Gd-DOTA não interrompeu a função das barreiras da orelha média e interna nem causou deficiência auditiva em ratos. Os marcadores biológicos inflamatórios críticos na orelha interna, incluindo a secreção de glicosaminoglicanos e ácido hialurônico e a expressão de CD44 e TLR2, não foram influenciados pela administração de LPNs. Não houve morte celular significativa associada à administração de LPNs. A injeção transtimpânica de LPNs é segura para o ouvido interno, e LPNs podem ser aplicados como uma matriz de distribuição de drogas na terapia clínica da perda auditiva neurossensorial.
Histórico
Os nanocarreadores de lipossomas (LPNs) são potencialmente o futuro da terapia do ouvido interno devido à sua alta capacidade de carga de drogas e captação eficiente no ouvido interno após uma administração intratimpânica minimamente invasiva [1,2,3,4]. A abordagem intratimpânica é bem aceita pelos otologistas como uma abordagem racional de administração de medicamentos direcionada porque evita o acúmulo desnecessário de agentes terapêuticos em regiões não direcionadas, o que tem sido uma estratégia anterior na clínica para o tratamento da doença de Meniere e perda auditiva neurossensorial súbita usando gentamicina e corticosteróides. O direcionamento molecular do modelo terapêutico na cóclea foi indicado pela administração intratimpânica de LPNs funcionalizados com peptídeos específicos [5]. Além disso, a entrega automática sustentada de LPNs para o ouvido interno através do ouvido médio foi alcançada usando um novo dispositivo composto de uma bomba osmótica e tubo de poliimida de alto desempenho [6]. Como a plataforma nanoterapêutica mais antiga da clínica, os LPNs eram seguros no tratamento de câncer, doenças infecciosas, inflamação, dor, etc. [7,8,9]. No entanto, a biocompatibilidade dos LPNs nas orelhas média e interna permanece desconhecida e precisa ser esclarecida antes que possam ser aplicados clinicamente em otologia.
A orelha é composta pelas orelhas externa, média e interna (fig. 1) que podem ser expostas aos LPNs após o parto intratimpânico. O ouvido médio é o local primário que é exposto aos LPNs em sua concentração mais alta, o ouvido interno é o local terapêutico e o órgão mais sensível aos agentes perigosos, e o canal auditivo externo tem o potencial de ser irritado por agentes que fluem do cavidade do ouvido médio. As barreiras biológicas são o primeiro sistema de defesa, limitando a biodisponibilidade dos agentes, e existem na pele, mucosa e estruturas perineurais. O sistema de barreira do ouvido interno desempenha um papel crítico na manutenção da homeostase iônica, essencial para a atividade fisiológica do ouvido interno. A alteração funcional dessas barreiras pode ser avaliada com precisão usando ressonância magnética com gadolínio (Gd-MRI). O comprometimento da função auditiva pode ser medido com precisão por meio da resposta auditiva do tronco encefálico (ABR). Portanto, a própria orelha (incluindo as orelhas externa, média e interna) serve como um excelente modelo para nanotoxicologia [10, 11].
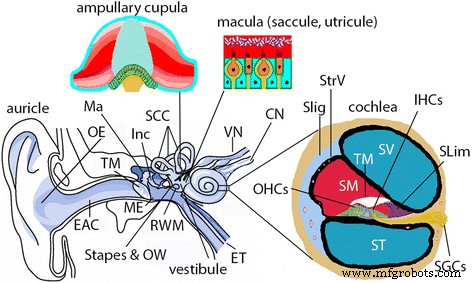
Ilustração da orelha de mamífero. O ouvido dos mamíferos (incluindo humanos e ratos) é composto pelas orelhas externa, média e interna. O ouvido externo ( OE ) é composto por aurícula e conduto auditivo externo ( EAC ) O ouvido médio ( EU ) é composto pela membrana timpânica ( TM ) e a cavidade que abriga a cadeia ossicular, incluindo o martelo ( Ma ), bigorna ( Inc ) e estribo. A cavidade do ouvido médio é uma extensão da nasofaringe através da trompa de Eustáquio ( ET ) e se comunica com o ouvido interno através da janela oval ( OW ) e membrana da janela redonda ( RWM ) A orelha interna é composta pela cóclea e sistema vestibular. A cóclea é o órgão sensorial da audição e tem três câmaras, ou seja, os compartimentos perilinfáticos da escala do tímpano ( ST ) e scala vestibuli ( SV ), e o compartimento endolinfático da escala média ( SM ) Na parede lateral da SM, há a estria vascular ( StrV ) e ligamento espiral ( SLig ) Na parte inferior do SM, existem órgãos de Cortis que contém células ciliadas internas ( IHCs ) e células ciliadas externas ( OHCs ), membrana tectorial ( TM ) e limbo espiral ( Slim ) As células ganglionares espirais ( SGCs ) disparar um potencial de ação correspondente à transdução mecânico-elétrica das células ciliadas e fornecer todas as entradas auditivas do cérebro. O sistema vestibular é responsável pelo equilíbrio e é composto por três canais semicirculares ( SCC ) e vestíbulo. A cúpula ampular dentro do SCC detecta acelerações rotacionais e a mácula dentro do sáculo e utrículo do vestíbulo detecta acelerações lineares. CN nervo coclear, SP proeminência espiral, VN nervo vestibular, VS vas spiralis. (adaptado de Zou J. Focal Drug Delivery in Inner Ear Therapy:in Focal Controlled Drug Delivery. Editores:Domb AJ e Khan W. Springer, Londres, Reino Unido. ISBN:978-1-4614-9433-1, 2014; p215- 224)
O ácido hialurônico (hialuronano) é um biopolímero polianiônico de ocorrência natural e é um componente primário da matriz extracelular na membrana basal. O ácido hialurônico é composto de ácido D-glucurônico e N-acetil-D-glucosamina, que estão ligados por meio de ligações glicosídicas alternadas β-1, 4 e β-1, 3. O acúmulo de ácido hialurônico pode contribuir para o aumento da permeabilidade e inflamação da microcirculação na lesão de reperfusão isquêmica renal [12]. O efeito ototóxico das nanopartículas de prata foi mostrado para ser correlacionado ao acúmulo de ácido hialurônico na cóclea de rato em nosso relatório anterior [11]. O ácido hialurônico se liga ao CD44 e ao receptor toll-like 2/4 (TLR2 / 4) no tecido e desencadeia reações biológicas [13, 14]. As atividades biológicas mediadas pelo CD44 após a ligação ao ácido hialurônico são principalmente através da interação com moléculas reguladoras e adaptadoras, como SRC quinases, Rho GTPases, VAV2, proteína ligada ao receptor do fator de crescimento 2-proteína de ligação associada 1 (GAB1), anquirina, e ezrin [15,16,17]. O CD44 também medeia o metabolismo do ácido hialurônico por meio de abordagens de captação e degradação celular, além de recrutar células T para locais inflamatórios e regular a lesão endotelial mediada por células T [18]. Foi relatado que a citotoxicidade para as células endoteliais do ouvido interno por anticorpos anti-células endoteliais pode desempenhar um papel em causar o dano da estria vascular na surdez neurossensorial súbita imunomediada [19]. A ativação do fator nuclear κB dependente de TLR2 estava supostamente envolvida na regulação positiva da proteína 1 quimiotática de monócitos induzida por Haemophilus influenzae nos fibrócitos do ligamento espiral do ouvido interno, que pode ser a etapa chave na disfunção do ouvido interno secundária à otite média crônica [ 20]. Se LPNs induzem comprometimento do ouvido interno após a administração do ouvido médio, a via de sinalização mediada por TLR2 deve ser o mecanismo importante.
Nosso objetivo foi avaliar a biocompatibilidade de LPNs na orelha interna após a injeção transtimpânica. As funções das barreiras biológicas na pele (canal auditivo externo), mucosa (cavidade da orelha média) e compartimentos da orelha interna foram medidas usando Gd-MRI em vários pontos de tempo. A função auditiva foi avaliada por meio da medida do ABR. Finalmente, as potenciais alterações histopatológicas foram analisadas medindo os acúmulos de glicosaminoglicanos e ácido hialurônico, a expressão de CD44 e TLR2 e a fragmentação de DNA na cóclea.
Resultados
LPNs não causaram alterações funcionais na cóclea de rato
No grupo controle positivo, sinal luminoso na perilinfa da cóclea (Coch) e vestibular (Colete) em ambos os lados (L, R) (Fig. 2a, b) indicando captação de Gd-DOTA. Após injeções transtimpânicas de nanopartículas de prata (AgNPs), as intensidades de sinal nos compartimentos perilinfáticos aumentaram significativamente, enquanto um sinal extremamente intenso também foi detectado na pele do conduto auditivo externo, mucosa da orelha média, indicando a captação aumentada de Gd-DOTA associada à administração de AgNP ( L na Fig. 2a, b) (Tabela 1). O sistema de avaliação foi, portanto, validado. No animal que recebeu injeção transtimpânica de LPN + Gd-DOTA, um sinal brilhante foi detectado na superfície da cadeia ossicular, escala vestibular, escala do tímpano e vestíbulo 3 h após a injeção, indicando distribuição óbvia de LPN nessas regiões (Fig. 2c , d). A intensidade do sinal na escala vestibular no giro basal foi visivelmente mais forte do que na escala do tímpano, sugerindo uma entrada eficiente de LPN através da janela oval no animal atual [21]. Às 6 h pós-injeção, as intensidades de sinal entre a escala vestibuli e escala do tímpano na volta basal tornaram-se semelhantes e toda a cóclea mostrou sinal quase homogêneo, mas houve alterações insignificantes no vestíbulo (Fig. 2e, f). Em animais que receberam injeções intravenosas de Gd-DOTA após injeção transtimpânica de LPNs em branco, ambos os lados exibiram intensidades de sinal semelhantes, exceto que havia sinais fortes no ouvido médio recebendo injeção transtimpânica de LPNs em branco, suspeitando de acúmulo de LPNs na superfície da cadeia ossicular (Fig. . 2g, h). O buraco negro na cadeia ossicular indicando a área oca do estribo (Fig. 2h). Intensidades de sinal iguais em ambos os lados sugeriram que a propriedade de transporte para Gd-DOTA das barreiras sangue-perilinfa em ambas as orelhas não mudou após a injeção transtimpânica de LPNs (Fig. 2g, h) (Tabela 1).
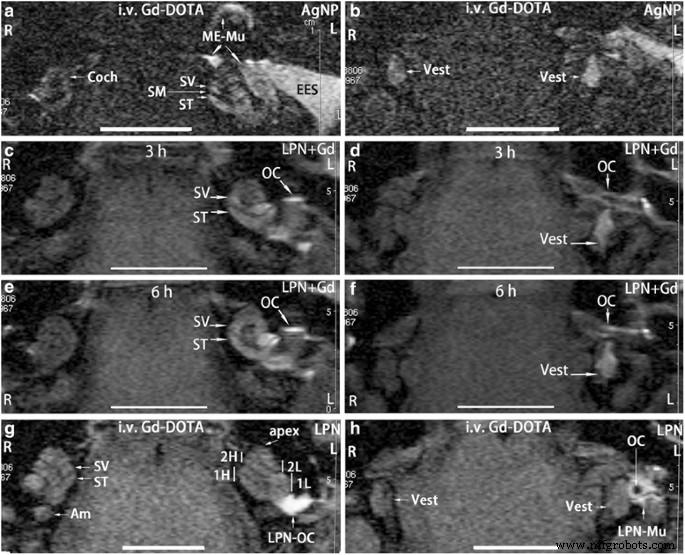
Ressonância magnética com gadolínio de orelha interna de rato após administração de nanocarreador de lipossoma (LPN). Em todos os animais, nanomateriais foram injetados na parede medial da cavidade do ouvido médio esquerdo. O controle positivo foi fotografado em ratos 2 h após a injeção intravenosa de Gd-DOTA secundária à injeção transtimpânica de nanopartículas de prata (AgNPs) 5 h antes ( a , b ) A distribuição dinâmica de LPNs nas orelhas média e interna foi mostrada em c , d , e , f por injeção transtimpânica de LPN contendo Gd-DOTA sem administração intravenosa de Gd-DOTA. O impacto de LPNs vazios na barreira biológica foi demonstrado por MRI 2 h após a injeção intravenosa de Gd-DOTA (i.v. Gd-DOTA) em ratos que receberam injeção transtimpânica de LPN 5 h antes ( g , h ) Sou ampular do canal semicircular posterior, Coch cóclea, EES pele da orelha externa, L orelha esquerda, LPN-Mu LPN na mucosa do ouvido médio, LPN-OC LPN na cadeia ossicular ( OC ), ME-Mu , mucosa do ouvido médio, R orelha direita, SM scala media , ST escala do tímpano, SV scala vestibuli, Colete vestíbulo, 1H espira basal superior da cóclea, 1L espira inferior basal da cóclea, 2H segunda espira superior da cóclea, 2L segunda volta inferior da cóclea. Barra de escala =5 mm
Nem LPN + Gd-DOTA nem LPNs causaram perda auditiva significativa, apresentada como uma mudança de limiar de ABR que foi medida usando estímulos de click e tone bursts nas frequências de 2, 4, 8, 16 e 32 kHz a 2, 4, e 7 dias após a administração, em comparação com as orelhas que receberam injeções transtimpânicas de água desionizada (dH 2 O) (Fig. 3).
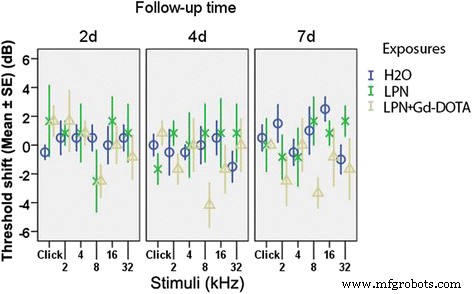
Impacto da injeção transtimpânica de nanocarreadores de lipossomas na função auditiva de ratos medida pela resposta auditiva do tronco encefálico. A perda auditiva foi expressa como mudanças de limiar. Houve diferença insignificante entre os grupos ( p > 0,05, ANOVA de uma via). n =6 em cada grupo. H2O injeção transtimpânica de água deionizada no grupo de controle negativo, LPN nanocarreador de lipossoma vazio, LPN + Gd-DOTA LPN contendo Gd-DOTA, 2d , 4d , e 7d 2, 4 e 7 dias após a injeção
LPNs não induziu acúmulo de glicosaminoglicano na cóclea de rato
A coloração com hematoxilina e eosina não demonstrou infiltração inflamatória de leucócitos e fibrina na cóclea de todos os animais analisados, incluindo estribo e janela oval por onde passam os LPNs (Fig. 4). A coloração de Schiff com ácido periódico demonstrou a existência de glicosaminoglicanos na parede óssea, limbo espiral, ligamento espiral, membrana tectorial, membrana de Reissner, lâmina espiral óssea e estria vascular na cóclea de animais que receberam injeções transtimpânicas de dH 2 O. Houve um aumento gradiente na intensidade do sinal do giro basal ao ápice, e a diferença foi significativa na estria vascular (figs. 5 e 6). O gradiente de sinal na cóclea não foi alterado nos animais que receberam injeção transtimpânica de LPNs e LPN + Gd-DOTA (Figs. 5 e 6).
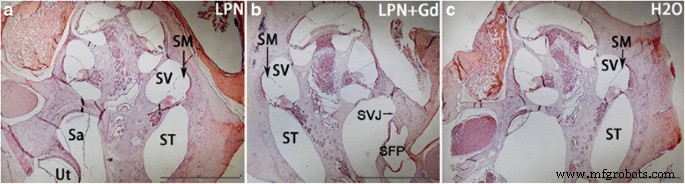
Coloração com hematoxilina-eosina de cócleas de ratos expostas a nanocarreadores de lipossomas. Não houve infiltrado inflamatório na cóclea que recebeu administrações de LPN ( a ), LPN + Gd ( b ) e H2O ( c ) Área circulada seleção indicada de região de interesse para medições de intensidade ( a ) LPN nanotransportador de lipossoma vazio, LPN + Gd LPN contendo Gd-DOTA. Sá sáculo, SFP plataforma do estribo, SVJ articulação estapediovestibular, SM scala media, ST escala do tímpano, SV scala vestibuli, Ut utrículo. Barra de escala =1 mm
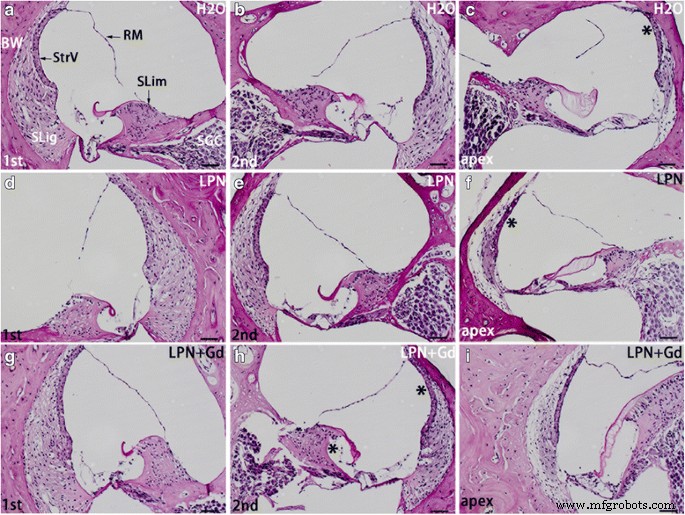
A secreção de glicosaminoglicano na cóclea de rato exposta a nanocarreadores de lipossoma foi detectada usando microscopia de luz de coloração de Schiff com ácido periódico. O limbo espiral ( SLim ) e parede óssea ( BW ) da cóclea mostrou a coloração mais intensa nos grupos de controle negativo ( H2O ) ( a - c ), nanocarreador de lipossoma vazio (LPN) ( d - f ), e LPN contendo Gd-DOTA ( LPN + Gd ) ( g - i ) A área de coloração com intensidades visivelmente mais altas foram indicadas por * em c , f , e h em comparação com a coluna da esquerda. RM Membrana de Reissner, SGC célula ganglionar espiral, SLig ligamento espiral, StrV estria vascular, 1ª volta basal, 2ª segundo turno. Barra de escala =50 μm
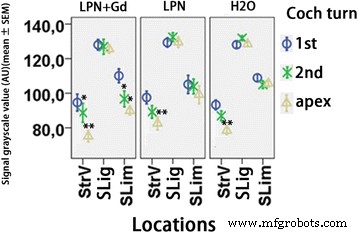
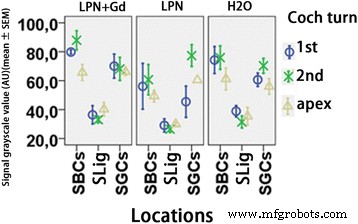
Quantificação da secreção de glicosaminoglicano na cóclea de rato exposta a nanocarreadores de lipossomas detectados usando a coloração de Schiff com ácido periódico. n =3 em cada grupo. AU unidade arbitrária, H2O controle negativo, LPN nanotransportador de lipossoma vazio, LPN + Gd LPN contendo Gd-DOTA, SLig ligamento espiral, Slim limbo espiral, StrV estria vascular, 1ª volta basal, 2ª segundo turno. * p <0,05, ** p <0,01 (ANOVA unilateral com teste LSD usado como análise post hoc)
Houve um pequeno impacto na secreção de ácido hialurônico na cóclea de rato por LPNs
Na cóclea de ratos que receberam injeções transtimpânicas de dH2O, a coloração positiva para ácido hialurônico foi detectada predominantemente nas células do gânglio espiral, células basais estriais, células do sulco externo e células endoteliais capilares, entre outras células (Fig. 7). As intensidades de sinal nos fibrócitos do ligamento espiral das voltas basal e segunda foram significativamente maiores do que no ápice. Essas diferenças tornaram-se insignificantes na cóclea de ratos com a aplicação de LPNs e LPN + Gd-DOTA, indicando que a secreção de ácido hialurônico pelos fibrócitos do ligamento espiral foi afetada pela administração de LPN (Fig. 8). LPN + Gd-DOTA também reduziu a coloração nos fibrócitos do ligamento espiral da volta basal. No entanto, não houve impacto na secreção de ácido hialurônico na maioria das células cocleares pela injeção transtimpânica de LPNs e LPN + Gd-DOTA (Figs. 7 e 8).
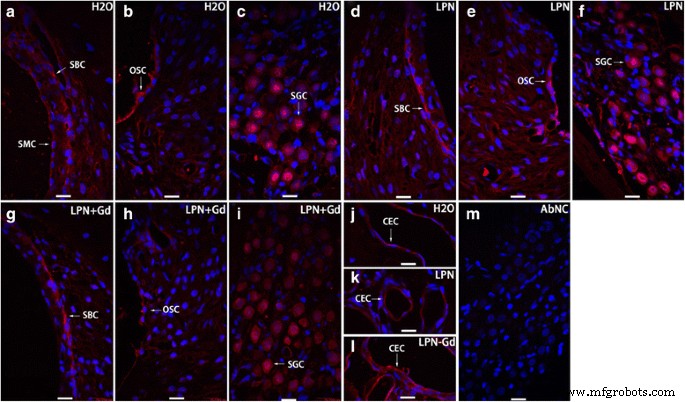
A secreção de ácido hialurônico na cóclea de rato exposta a nanocarreadores de lipossomas foi detectada com microscopia confocal imunofluorescente. A coloração positiva foi encontrada na célula basal da estria ( SBC ), célula do sulco externo ( OSC ), célula ganglionar espiral ( SGC ), e célula endotelial capilar ( CEC ) de modíolo de grupos de controle negativo ( H2O ) ( a - c , j ), nanocarreadores de lipossomas vazios ( LPN ) ( d - f , k ), e LPN contendo Gd-DOTA ( LPN + Gd ) ( g - i , l ) Não houve coloração no controle negativo omitido do anticorpo ( AbNC ) ( m ) CEC célula endotelial capilar, ISC célula do sulco interno, SBC célula basal da estria, SL-I fibrócito de ligamento espiral tipo I, SLSF fibrócito satélite de limbo espiral, SMC célula marginal da estria vascular. Barra de escala =16 μm
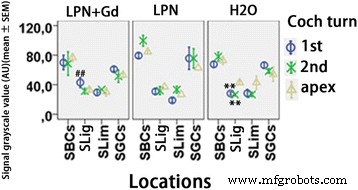
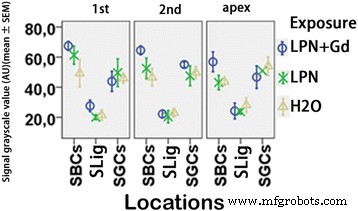
Quantificação da secreção de ácido hialurônico em cóclea de rato exposta a nanocarreadores de lipossomas detectados por microscopia confocal imunofluorescente. n =3 em cada grupo. AU unidade arbitrária, H2O controle negativo, LPN nanotransportador de lipossoma vazio, LPN + Gd LPN contendo Gd-DOTA, SBCs células basais da estria, SGCs células ganglionares espirais, SLig , ligamento espiral, Slim limbo espiral, 1º volta basal, 2ª segundo turno. ** p <0,01 (em comparação com o ápice), ## p <0,01 (comparando com os grupos LPN e H2O da volta basal) (ANOVA de uma via com teste LSD usado como análise post hoc)
LPNs não alterou a população de células CD44 na cóclea de rato
Na cóclea exposta ao dH 2 O, as células intermediárias estriais, células basais estriais, fibrócitos do ligamento espiral, células do gânglio espiral, células de Deiters no órgão de Corti e células endoteliais capilares no modíolo e ligamento espiral mostraram coloração intensiva para CD44. Houve uma diferença insignificante nas intensidades de sinal entre as voltas cocleares. A população CD44-positiva e a intensidade de expressão não foram afetadas pela injeção transtimpânica de LPN + Gd-DOTA ou LPNs (Figs. 9 e 10).
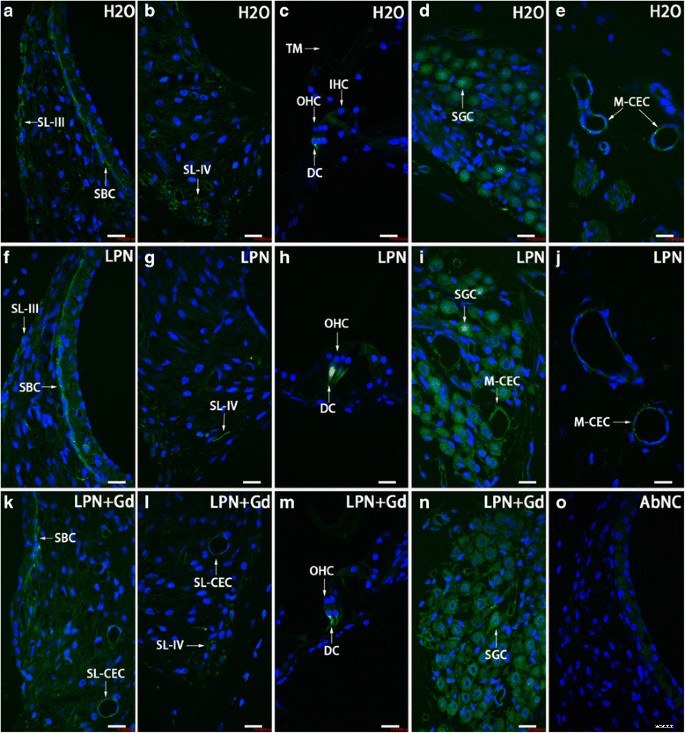
Distribuição de células CD44-positivas na cóclea de rato exposta ao nanocarreador de lipossoma demonstrada usando microscopia confocal imunofluorescente. As células positivas para CD44 foram detectadas principalmente na célula basal da estria ( SBC ), ligamento espiral ( SL ), Células de Dieter ( DC ), célula ganglionar espiral ( SGC ), e célula endotelial capilar ( CEC ) nos grupos de controle negativo ( H2O ) ( a - e ), nanocarreadores de lipossomas vazios ( LPN ) ( f - j ), e LPN contendo Gd-DOTA ( LPN + Gd ) ( k - n ) Não houve coloração no controle negativo omitido do anticorpo ( AbNC ) ( o ) IHC células ciliadas internas, M-CEC célula endotelial capilar em modíolo, SL-CEC célula endotelial capilar no ligamento espiral:células do gânglio espiral, SL-III fibrócito do ligamento espiral tipo III, SL-IV fibrócito do ligamento espiral tipo IV, TM membrana tectorial, OHC células ciliadas externas. Barra de escala =16 μm
Quantificação do nível de proteína CD44 em cóclea de rato exposta a nanocarreadores de lipossomas detectados por microscopia confocal imunofluorescente. Houve diferença insignificante entre os grupos ( p > 0,05, ANOVA de uma via). n =3 em cada grupo. n =3 em cada grupo. AU unidade arbitrária, H2O controle negativo, LPN nanotransportador de lipossoma vazio, LPN + Gd LPN contendo Gd-DOTA, SBCs células basais da estria, SGCs células ganglionares espirais, SLig ligamento espiral, 1º volta basal, 2ª segunda volta
LPNs não alterou a expressão de TLR2 na cóclea de rato
Na cóclea exposta ao dH2O, as células basais estriais, fibrócitos do ligamento espiral, células da raiz, células do gânglio espiral, células pilares do órgão de Corti e células endoteliais capilares no modíolo mostraram coloração intensiva para TLR2. Houve uma diferença insignificante nas intensidades de sinal entre as voltas cocleares. A população TLR2-positiva e a intensidade de expressão não foram afetadas pela injeção transtimpânica de LPN + Gd-DOTA ou LPNs (Figs. 11 e 12).
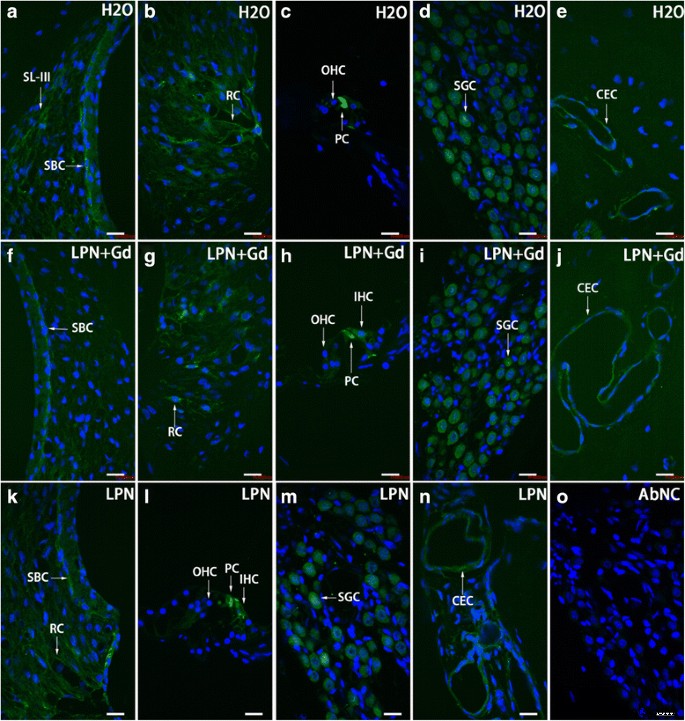
Distribuição de células positivas para TLR2 na cóclea de rato exposta ao nanocarreador de lipossoma demonstrada usando microscopia confocal imunofluorescente. As células positivas para TLR2 foram detectadas principalmente na célula basal da estria ( SBC ), ligamento espiral ( SL ), célula raiz ( RC ), célula pilar ( PC ), célula ganglionar espiral ( SGC ), e célula endotelial capilar ( CEC ) nos grupos de controle negativo ( H2O ) ( a - e ), nanocarreadores de lipossomas vazios ( LPN ) ( k - n ), e LPN contendo Gd-DOTA ( LPN + Gd ) ( f - j ) Não houve coloração no controle negativo omitido pelo anticorpo ( AbNC ) ( o ) IHC células ciliadas internas, SL-III fibrócito de ligamento espiral tipo III, OHC células ciliadas externas. Barra de escala =16 μm
Quantificação do nível de proteína TLR2 em cóclea de rato exposta a nanocarreadores de lipossomas detectados por microscopia confocal imunofluorescente. Houve diferença insignificante entre os grupos ( p > 0,05, ANOVA de uma via). n =3 em cada grupo. AU unidade arbitrária, H2O controle negativo, LPN nanotransportador de lipossoma vazio, LPN + Gd LPN contendo Gd-DOTA, SBCs células basais da estria, SGCs células ganglionares espirais, SLig ligamento espiral, 1º volta basal, 2ª segunda volta
LPNs não causou morte celular na cóclea de rato
Havia células apoptóticas esparsas que são distribuídas aleatoriamente na cóclea de ratos não tratados. Surpreendentemente, havia células apoptóticas abundantes na platina do estribo e no nicho da janela oval. Não houve impacto na quantidade e padrão de distribuição de células apoptóticas pela administração de LPNs e LPN + Gd-DOTA (Fig. 13).
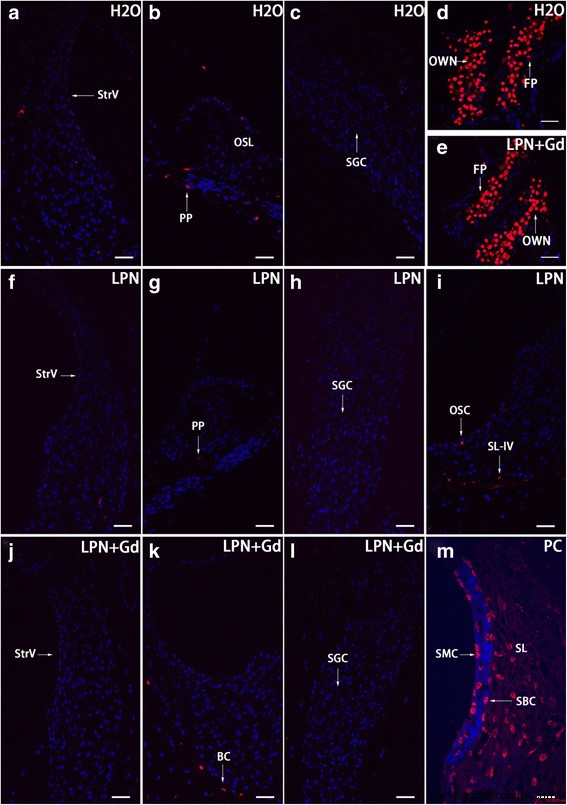
Apoptose na cóclea de rato exposta a nanocarreadores de lipossoma demonstrada usando microscopia confocal de coloração TUNEL. Células apoptóticas foram detectadas esparsamente nas células cocleares de ratos em grupos de controle negativo ( H2O ) ( a - c ), nanocarreadores de lipossomas vazios ( LPN ) ( f - i ), e LPN contendo Gd-DOTA ( LPN + Gd ) ( j - l ) Havia células apoptóticas abundantes na platina do estribo ( FP ) e nicho de janela oval ( PRÓPRIO ) de ambos os grupos ( d , e ) Em um controle positivo (PC), coloração TUNEL abundante foi detectada nos fibrócitos do ligamento espiral ( SL ), células basais da estria ( SBC ), e células estria margina ( SMC ) PP processo periférico da célula ganglionar espiral ( SGC ), OC osteócito, OSC célula do sulco externo, SL-IV tipo IV de fibrócitos do ligamento espiral, SLim limbo espiral, StrV estria vascular. Barra de escala a - l =32 μm, m =16 μm
Discussão
LPNs entraram no ouvido interno de forma eficiente após a injeção transtimpânica demonstrada por ressonância magnética usando Gd-DOTA como miméticos de drogas que foram encapsulados dentro dos LPNs (Fig. 2c-f). Embora um estudo anterior tenha mostrado que a janela redonda era o principal caminho dos LPNs para entrar no interior [6], a presente observação mostrou que o caminho da janela oval era mais eficiente do que a janela redonda para transportar os LPNs do ouvido médio para o interior orelha. Este resultado sugeriu que ambas as vias são importantes na carga da orelha interna de LPNs após a administração da parede medial da orelha média direcionada. Usando o método in vivo mais eficiente de ressonância magnética do ouvido interno com gadolínio para avaliar a barreira biológica e ABR específico de frequência para avaliar a função auditiva, o presente estudo demonstrou que a injeção transtimpânica de LPNs e LPN + Gd-DOTA não interrompeu a função de as barreiras do ouvido interno nem causaram deficiência auditiva em ratos. Ao analisar os marcadores biológicos inflamatórios críticos previamente demonstrados [11, 22], LPNs e LPN + Gd-DOTA não induziram a resposta inflamatória na cóclea. Embora a membrana da janela redonda não tenha sido avaliada, a ausência de inflamação no estribo e na janela oval descartou uma reação inflamatória óbvia na membrana da janela redonda, uma vez que o presente estudo demonstrou que a via da janela oval foi superior à abordagem da janela redonda para LPNs.
A ressonância magnética do ouvido interno após a injeção intravenosa de quelato de gadolínio é capaz de detectar a ruptura mediada pelo estresse oxidativo nas barreiras da perilinfa e da endolinfa sanguínea induzida por toxinas mitocondriais [23]. Foi relatado que AgNPs causam comprometimento celular por meio da geração de espécies reativas de oxigênio (ROS) e a ativação de quinases do terminal amino de Jun (JNK), levando à liberação de citocromo C no citosol e à translocação de Bax para a mitocôndria [24 ] Na injeção transtimpânica, os AgNPs entraram na orelha interna e induziram mudanças de permeabilidade nas barreiras biológicas da orelha interna do rato [11, 25]. LPNs também entraram no ouvido interno do rato após a injeção transtimpânica em um padrão dependente do tamanho, e os LPNs de 95 nm de diâmetro mostraram a maior eficácia na passagem através das barreiras do ouvido interno médio [3]. No presente estudo, o tamanho médio dos LPNs foi de 100 a 115 nm, que era um pouco maior do que o tamanho mais eficiente. LPN + Gd-DOTA mostrou que esse tamanho de LPNs entrou na orelha interna, o que está de acordo com o relato anterior [3]. No entanto, a entrada de LPNs no ouvido interno não causou alterações de permeabilidade nas barreiras sangue-perilinfa e sangue-endolinfa. Este resultado sugere que os LPNs são seguros para o ouvido interno. Os resultados do ABR indicando uma função auditiva normal apoiaram o resultado da ressonância magnética.
Houve associação entre secreção de ácido hialurônico e alteração da permeabilidade e inflamação da microcirculação na lesão de reperfusão isquêmica renal [12]. The previous study also showed that AgNPs caused the accumulation of hyaluronic acid in the rat cochlea [11]. CD44 and toll-like receptor 2/4 (TLR2/4) work as receptors of hyaluronic acid and trigger biological reactions [13, 14]. CD44 also mediates the metabolism of hyaluronic acid through cellular uptake and degradation in addition to recruiting T cells to inflammatory sites and regulating T cell-mediated endothelial injury [18]. In the present study, hyaluronic acid, CD44, and TLR2 were detected in the rat cochlea. LPN + Gd-DOTA reduced the secretion of hyaluronic acid in the spiral ligament fibrocytes. The expressions of CD44 and TLR2 were not changed after the transtympanic injection of either LPNs or LPN + Gd-DOTA. The total glycosaminoglycan, which contains hyaluronic acid in the cochlea was not affected by the administrations of LPNs and LPN + Gd-DOTA. The impact of LPNs on the hyaluronic acid distribution in rat cochlea did not cause either permeability change or hearing loss, indicating that the modification is unharmful. It was reported that macrophages undergo phenotypic changes dependent on molecular weight of hyaluronan that correspond to either pro-inflammatory response for low molecular weight hyaluronic acid or anti-inflammatory response for high molecular weight hyaluronic acid [26]. The observed minor changes of hyaluronic acid distribution in the cochlea might have high molecular weight and anti-inflammatory function. Therefore, there was no hint of an inflammatory reaction in the rat cochlea.
The observed apoptosis in the stapes footplate cells might be normal biological activity. A balance between survival and apoptosis in the stapes footplate cells was reportedly as necessary to inactivate the otosclerosis [27]. Administration of LPN + Gd-DOTA did not affect apoptosis in the rat stapes.
Conclusions
The present study demonstrated that the transtympanic injection of liposome nanocarriers neither impaired the biological barriers of the inner ear nor caused hearing loss in the rats. The critical inflammatory mechanism was not activated by the administration of liposome nanocarriers, either. The results suggested that transtympanic injection of liposome nanocarrier is safe for the cochlea of rat.
Methods
Materials
Sphingosine (Sph), 1-stearoyl-2-oleoyl-sn-glycero-3-phosphocholine (SOPC), and 1, 2-distearoyl-sn-glycero-3-phosphoethanolamine-N-[methoxy(polyethyleneglycol)-2000] (ammonium salt) [DSPE-PEG-2000] were purchased from Avanti polar lipids (Alabaster, USA). DiI (Vybrant DiI cell-labeling solution, 1 mM in solvent) and N-(6-tetramethylrhodaminethiocarbamoyl)-1,2-dihexadecanoyl-sn-glycero-3-hosphoethanolamine, triethylammonium salt (TRITC-DHPE) were purchased from Thermo Fisher Scientific (Waltham, USA). Gd-DOTA (DOTAREM) was from Guerbet, Cedex, France. Hepes and EDTA were from Sigma. The purity of the lipids was evaluated using thin-layer chromatography on silicic acid-coated plates (Merck, Darmstadt, Germany) developed with a chloroform/methanol/water mixture (65:25:4, v/v/v). An examination of the plates after iodine staining and, when appropriate, upon UV illumination revealed no impurities. The lipid concentrations were determined gravimetrically with SuperG (Kibron, Espoo, Finland); a high-precision microbalance. The polyvinylpyrrolidone-stabilized AgNPs were supplied by Colorobbia (Firenze, Italy). The AgNPs were dispersed in deionized water (370.7 mM), and scanning electron microscopy showed that the AgNPs are spheroids with a particle size of around 100 nm. Dynamic light scattering (DLS) showed a mean hydrodynamic size of 117 ± 24 nm and a mean zeta potential of −20 ± 9 mV.
In the visualization of nanocarrier uptake in the inner ear and the evaluation of biological barrier function, 5 male Sprague Dawley rats, weighing between 334 and 348 g, were provided by the Biomedicum Helsinki, Laboratory Animal Centre, University of Helsinki, Finland (this is the defined animal center that provides animals for MRI experiments in Biomedicum); in the ABR and histological studies, 18 Sprague Dawley rats, weighing between 300 and 400 g, were provided by the Experimental Animal Unit of the University of Tampere School of Medicine in Finland. Animal assignments in each study were shown in Table 2. All animal experiments were approved by the Ethical Committee of University of Tampere (permission number:ESAVI/3033/04.10.03/2011). Animal care and experimental procedures were conducted in accordance with European legislation. Animals in the Gd-MRI study were anesthetized with isoflurane with 5% isoflurane–oxygen mixture (flow-rate 1.0 L/min) for induction and 3% for maintenance via a facemask. Animals for the ABR and histological studies were anesthetized with a mixture of 0.5 mg/kg medetomidine hydrochloride (Domitor ® , Orion, Espoo, Finland) and 75 mg/kg ketamine hydrochloride (Ketalar ® , Pfizer, Helsinki, Finland) via intraperitoneal injection followed by an intramuscular injection of enrofloxacin (Baytril ® vet, Orion, Turku, Finland) at a dose of 10 mg/kg to prevent potential infection. The animal’s eyes were protected by Viscotears® (Novartis Healthcare A/S, Copenhagen, Denmark).
Preparation and Characterization of LPNs with and without Gd
Preparation of Gd-containing LPNs
LPNs of unilamellar vesicles with an apparent hydrodynamic particle diameter (Z av ) of 110 ± 15 nm that contain Gd-DOTA were prepared according to the previously published method [6]. A concentration of 1 mM Gd-DOTA-containing LPN (LPN + Gd-DOTA) refers to 1 mM liposomes encapsulating 500 mmol/L of Gd-DOTA.
Preparation of Blank LPNs
Blank LPNs of unilamellar vesicles with Z av of 115 ± 10 nm were prepared according to a previous publication [6].
Administration of LPNs
Under general anesthesia with isoflurane with 5% isoflurane–oxygen mixture (flow-rate 1.0 L/min), 50 μl of either LPNs or LPN + Gd-DOTA were injected into the left middle ear cavity through the tympanic membrane penetration under an operating microscope (OPMI1-F, Carl Zeiss, Jena, Germany). The same amount of deionized water (H2 O) was injected transtympanically in rats that were assigned to the negative group. After the injection, the animals were kept in the lateral position with the injected ear oriented upward for 15 min before further measurements.
Evaluation on Biological Barrier Function Using Gd-MRI
One animal receiving transtympanic injection of LPN + Gd-DOTA was selected to demonstrate distributions of LPN in the inner ear using MRI without contrast agent. Two animals receiving transtympanic injection of blank LPNs were engaged in MRI study for evaluation of the biological barrier function. Two animals receiving transtympanic injection of AgNPs (370.7 mM, 40 μl) were used as positive control. The contralateral ear without any injection was used as negative control in all studies. A 4.7T MR scanner with a bore diameter of 155 mm (PharmaScan, Bruker BioSpin, Ettlingen, Germany) was utilized. The maximum gradient strength was 300 mT/m with an 80-μs rise time. A gadolinium-tetraazacyclododecane-tetraacetic acid (Gd-DOTA, 500 mM, DOTAREM, Guerbet, Cedex, France) solution was injected into the tail vein (0.725 mM/kg) 2 h before the MRI measurements. The imaging protocol and rapid acquisition with relaxation enhancement (RARE) sequences were applied according to a previous publication [10]. MRI scanning commenced at several time points after the transtympanic injection. The first MRI time of around 5 h was determined by taking the penetration time of liposome nanoparticles from the middle ear to the inner ear as a reference [1, 3, 6]. The final imaging time of 8 d was selected according to the course of potential acute inflammation and the availability of the scanner. ParaVision PV 4.0 (Bruker, MA, USA) software was used for the post-processing and quantification of MR images.
ABR Measurement
The auditory function of animals receiving injections of both blank and Gd-containing LPNs were evaluated using ABR measurements using BioSig32 (Tucker Davis Technologies, FL, USA) in a custom made, soundproof chamber. The ABR thresholds upon click and tone burst stimuli were recorded before and at a certain time point post-administration of LPNs. The first ABR measurement was followed on 2 days post-administration of AgNPs, allowing the animals to recover from the general anesthesia during the injection and to ensure the injected solution to be entirely cleared from the middle ear cavity. The second follow-up time of 4 days post-injection was chosen because it is close to the peak time of potential mitochondrial impairment-induced cell death in the cochlea [22]. The third follow-up time of 7 days is the time point when temporary threshold shifts remained significantly approved in an animal model of mitochondrial toxin-induced hearing loss [28]. The ABR recording procedure followed the previous report [11].
Glycosaminoglycan Staining in Rat Cochlea After Administration of LPNs
Hematoxylin-eosin staining to assess potential inflammatory infiltration and periodic acid Schiff’s staining to evaluate potential glycosaminoglycan accumulation in the cochlea after administration of LPNs were performed according to a previous publication after ABR measurements over 7 days [11] The slices were observed and digital images were acquired under a light microscope (Leica DM2000 microscope equipped with an Olympus DP25 camera) for further analysis.
Immunofluorescence Staining for Hyaluronic Acid and Receptors
Immunofluorescence staining for hyaluronic acid, CD44, and TLR2 were performed according to a previous publication after ABR measurements over 7 days [11, 21].
Cell Death Detection
Potential nuclear DNA fragmentation in the cochlea was investigated using terminal transferase (TdT) to label the free 3′OH breaks in the DNA strands of apoptotic cells with TMR-dUTP (TUNEL staining) following the reported procedure [11]. Slices exposed to recombinant DNase I (Fermentas, Vantaa, Finland, 100 U/ml in 50 mM Tris/HCl, pH 7.5, 1 mg/ml bovine serum albumin) at 37 °C for 10 min, which induced DNA strand breaks prior to the labeling procedures, were utilized as positive controls. The samples were observed under a confocal microscope.
Confocal Microscopy
The samples were observed under a Nikon inverted microscope (ECLIPSE Ti) combined with an Andor confocal system installed with Andor iQ 2.8 software (Andor Technology, Belfast, UK). The excitation lasers were 488 nm (green excitation) and 568 nm (red excitation) from an Andor laser combiner system, and the corresponding emission filters were 525/50 (Alexa Fluor-488) and 607/45 nm (Cy TM 3 and TMR Red). DAPI was excited with light at 405 nm generated from a light-emitting diode and was detected using a 450–465 nm filter.
Analysis and Statistics
ImageJ (1.45S, National Institutes of Health, Bethesda, USA) software was used for signal intensity measurements. For light microscopy of periodic acid Schiff’s staining, the region of interests (ROIs) including spiral ligament, spiral limbus, and stria vascularis were selected using freehand selections button. The “measure” function was used to obtain the mean gray scale value of the ROI, which was inversely correlated with the staining intensity. For confocal microscopy of immunofluorescence staining, the ROIs including stria basal cells, spiral ganglion cells, spiral ligament, and spiral limbus were extracted using photoshop CS6 (version 13.0, Adobe Systems Software Ireland Ltd, Dublin, Ireland) program and were imported into ImageJ program. The images were split into individual channel, and the green (corresponded to CD44 and TLR2) and red (corresponded to hyaluronic acid) channels were selected for further quantifications. The “Threshold” was adjusted using the “set” button in the “Image” menu, and “Limit to Threshold” option should be selected and “Direct to” should be defined to the corresponding channel in the “Analyze” menu. Then the gray scale value, which was inversely correlated with the staining intensity, was obtained using the “Measure” function in the same menu.
Statistical analyses were performed using the IBM ® SPSS ® Statistics Version 20 software package (SPSS Inc., Chicago, USA). A one-way ANOVA and Kruskal-Wallis test were used to compare ABR threshold shifts and signal intensities (grayscale) of staining for glycosaminoglycan and hyaluronic acid secretions, and TLR2 and CD44 staining between the LPN injected-ear and saline injected-ear groups in the different cochlear structures among various turns. Least significant difference (LSD) test was used as post hoc analysis. Higher numbers in the grayscale analysis correlate with lower signal intensities of the staining. p < 0.05 was accepted as statistically significant.
Abreviações
- ABR:
-
Auditory brainstem response
- AgNPs:
-
Silver nanoparticles
- dH2 O:
-
Água desionizada
- GAB1:
-
Growth factor receptor-bound protein 2-associated-binding protein 1
- Gd-DOTA:
-
Gadolinium-tetra-azacyclo-dodecane-tetra-acetic acid (DOTAREM)
- Gd-MRI:
-
Gadolinium-enhanced magnetic resonance imaging
- JNK:
-
Jun amino-terminal kinases
- LPN + Gd-DOTA:
-
Gd-DOTA-containing LPNs
- LPNs:
-
Liposome nanocarriers
- ROI:
-
Region of interests
- ROS:
-
Reactive oxygen species
- TLR:
-
Toll-like receptor
Transistores de efeito de campo Nanoflake SnSe multicamadas com contatos óhmicos de baixa resistência
Rota de eletrofiação em uma etapa de nanofibras de Rutilo TiO2 modificadas com SrTiO3 e suas propriedades fotocatalíticas
Nanomateriais
- Em sintonia com o coração de um átomo de cobre
- Detectando o magnetismo do núcleo de um átomo único
- Carbono oceânico gerado na escala atômica
- Após o crescimento, trazendo sistemas de calibração outlier para dentro da multinacional
- Após 15 segundos, o cérebro esquece as informações completamente
- Google encontra seu Glass após dois anos no escuro
- O Projeto da Camada de Emissão para Multiplicadores de Elétrons
- Amazon-Whole Foods:Dois anos após o negócio
- Após o protótipo:a próxima fase do processo de design
- Vida útil da bomba:após a reconstrução