


Nanoscopia baseada em microscopia de força atômica de células-tronco humanas derivadas de adiposo diferenciando condrogenicamente:nanoestrutura e expressão β1 de integrina
Resumo
A integrina β1 é conhecida por estar envolvida na diferenciação, migração, proliferação, reparo de feridas, desenvolvimento de tecidos e organogênese. A fim de analisar a probabilidade de ligação entre o ligante integrina β1 e o agrupamento de receptores de diferenciação 29 (CD29), microscopia de força atômica (AFM) foi usada para detectar receptores acoplados a integrina β1 nativa na superfície de células-tronco humanas derivadas de adiposo (hADSc) . A probabilidade de ligação da interação ligante-receptor da integrina β1 foi investigada por pontas funcionalizadas com integrina β1 em hADSc durante a diferenciação condrogênica inicial no nível de cultura de células bidimensionais. A morfologia celular e a ultraestrutura de hADSc foram medidas por AFM, que demonstrou que as células fusiformes longas tornaram-se células poligonais com relações comprimento / largura diminuídas e rugosidade aumentada durante a indução condrogênica. A ligação do ligante β1 da integrina e dos receptores CD29 foi detectada por pontas funcionalizadas com β1 para hADSc vivo. Um total de 1200 curvas foram registradas em 0, 6 e 12 dias de indução condrogênica. As forças de ruptura médias foram, respectivamente, 61,8 ± 22,2 pN, 60 ± 20,2 pN e 67,2 ± 22,0 pN. Os eventos de ruptura foram 19,58 ± 1,74%, 28,03 ± 2,05% e 33,4 ± 1,89%, respectivamente, o que demonstrou que a probabilidade de ligação foi aumentada entre o ligante β1 da integrina e os receptores na superfície do hADSc durante a indução condrogênica. A integrina β1 e a via de sinalização β-catenina / SOX foram correlacionadas durante a diferenciação condrogênica. Os resultados desta investigação implicam que AFM oferece uma visão cinética e visual sobre as mudanças na ligação do ligante β1 da integrina β1-receptor CD29 em hADSc durante a condrogênese. Mudanças na morfologia celular, ultraestrutura da membrana e probabilidade de ligação ligante-receptor transmembrana demonstraram ser marcadores úteis para avaliação do processo de diferenciação condrogênica.
Histórico
A osteoartrite (OA) é uma doença articular degenerativa comum em idosos [1], com OA degenerativa caracterizada pela destruição progressiva da cartilagem articular. A cartilagem é altamente organizada, sem vasos sanguíneos, nervos ou tecido linfático [2]. A matriz extracelular (MEC) é composta principalmente por colágeno II e glicoproteína, e é muito importante para a homeostase da cartilagem. Como a cartilagem é avascular, sua capacidade de autorrenovação é limitada. Embora os tratamentos de OA (cirúrgicos e não cirúrgicos) possam aliviar rapidamente os sintomas do paciente com OA, especialmente a dor, eles não podem restaurar a estrutura e função normais da cartilagem articular [3]. No futuro, o tratamento provavelmente incluirá engenharia de tecidos com células-tronco e estruturas para reparar defeitos e cartilagem articular degenerativa [4]. As células-tronco mesenquimais são células estromais multipotentes que apresentam potencial osteogênico, adipogênico, condrogênico e miogênico, dependendo das combinações de fatores de crescimento [5]. A análise da diferenciação de células-tronco mesenquimais mostrou Wnt / β-catenina, alvo mamífero da rapamicina (mTOR), fosfoinositídeo 3-quinase (PI3K) e outras vias para desempenhar papéis importantes na diferenciação [6,7,8]. No entanto, o mecanismo subjacente pelo qual a diferenciação condrogênica é induzida permanece indefinido. Isso é particularmente verdadeiro para o mecanismo pelo qual os sinais extracelulares ativam as vias de sinalização intracelular. Descobrimos que a integrina β1 sofre alterações durante a diferenciação condrogênica. Portanto, hipotetizamos que a integrina β1 pode desempenhar um papel importante na diferenciação condrogênica de células-tronco derivadas de adiposo humano (hADSc) devido ao seu envolvimento em várias vias de sinalização de diferenciação de tecido. Nesta investigação, o foco foi na via de sinalização Wnt / β-catenina.
Numerosos estudos têm mostrado que as interações entre as células e o ambiente extracelular são reguladas por proteínas transmembrana, em particular, membros da família das integrinas [9]. As integrinas são compostas por glicoproteínas transmembranares heterodiméricas de cadeias α e β não ligadas covalentemente [10]. Teoricamente, existem 64 integrinas conhecidas, das quais apenas 24 foram encontradas. As integrinas desempenham papéis vitais na adesão célula-célula, adesão ECM-célula, sinalização celular e organização do citoesqueleto de actina [11]. O ECM desempenha um papel importante na homeostase do tecido, sendo que o ECM regula as integrinas. As integrinas medeiam muitos processos fundamentais, incluindo adesão celular, migração, proliferação, diferenciação, morte celular, reparo de feridas, desenvolvimento de tecido e organogênese. Durante a diferenciação condrogênica das células-tronco mesenquimais, a expressão da integrina β1 está conectada à via de sinalização SOX e ao colágeno II. O foco desta investigação foi no dímero β1 da integrina, uma vez que é o dímero β mais proeminente entre os heterodímeros da cartilagem e é conhecido por interagir com muitos dímeros α diferentes [12]. O cluster de diferenciação 29 (CD29) é uma subunidade β1 da integrina associada a receptores de antígenos muito tardios, expressa em quase todas as células e tipos de tecido.
Aqui, a microscopia de força atômica (AFM) foi usada para nos ajudar a medir as mudanças durante a diferenciação condrogênica de hADSc. Como um tipo de microscopia de varredura de alta resolução, o AFM ofereceu uma nova oportunidade para detectar a morfologia e a membrana celular de células individuais em fluido em nanoescala. Enquanto isso, o sistema de espectroscopia de força de molécula única (SMFS) combinado por microscopia de força atômica (AFM) foi usado para medir a ligação ligante-receptor nas células vivas. O sistema de SMFS foi mais sensível às mudanças de receptores na membrana celular, e as imagens da força de ligação foram visualizadas. Neste trabalho, a ligação do ligante β1 da integrina β1 foi testada por pontas AFM funcionalizadas com integrina β1. Aplicando AFM, a diferenciação condrogênica foi encontrada para alterar a forma da célula hADSc e aumentar a rugosidade celular. Este aplicativo forneceu um método para avaliar a diferenciação condrogênica por medição direta das interações ligante β1 da integrina-receptor e alteração da ultraestrutura da superfície celular, melhorando a investigação da superfície celular e a triagem de uma forma visualizada. A diferenciação condrogênica altera a composição e estrutura da membrana, bem como as interações citoesqueléticas intracelulares. Essas mudanças na morfologia celular, ultraestrutura e ligação ligante-receptor transmembrana servem como marcadores úteis para a avaliação dos mecanismos de diferenciação condrogênica.
Métodos
Cultura de células e reagentes
Para esta investigação, células foram isoladas de três pacientes cirúrgicos (idade média de 20 anos) conforme descrito anteriormente [13]. Consentimentos informados foram obtidos de todos os pacientes. A aprovação ética para este estudo foi obtida do First Affiliated Hospital of Jinan University (formulário de suplemento). As células foram mantidas em meio basal, que incluiu meio de Eagle modificado por Dulbecco com baixo teor de glicose (DMEM, Life Technologies, CA, EUA) suplementado com 10% de soro fetal bovino inativado por calor (FBS, Life Technologies, CA, EUA), 100 unidades / ml de penicilina (Life Technologies, CA, EUA), estreptomicina 100 μg / ml (Life Technologies, CA, EUA), piruvato de sódio 0,11 mg / ml (Life Technologies, CA, EUA) e L-glutamina (Life Technologies, CA, EUA). As células foram mantidas a 37 ° C em uma incubadora umidificada contendo 5% de CO 2 com meio trocado a cada 3 dias.
Diferenciação in vitro
Para indução condrogênica, hADSc de quarta a oitava passagem foram semeadas em uma densidade celular alta (2 × 105/10 ml) e cultivadas em meio condrogênico contendo DMEM / F12 suplementado com 1% de FBS, 1% de Insulina-Transferrina-Selênio ( ITS) + suplemento (Cyagen, Guangzhou, China), fator de crescimento transformador beta1 de 10 ng / ml (TGF-β1) (Peprotech, Rocky Hill, New Jersey, EUA), fator de crescimento semelhante à insulina-1 de 100 ng / ml ( IGF-1) (Peprotech, Peprotech, Rocky Hill, New Jersey, EUA), dexametasona 10-7 M (Sigma, St. Louis, MO, EUA) e ácido ascórbico 50 μg / ml (Sigma, St. Louis, MO , EUA). O meio foi trocado a cada 2 dias com TGF-β1 e IGF-1 recém-adicionado. A condrogênese foi avaliada por coloração com azul alcian e azul de toluidina.
Para induzir a diferenciação osteogênica e adipogênica, as células da quarta à oitava passagem foram tratadas com o meio osteogênico e adipogênico por 2 semanas, respectivamente. O meio osteogênico consistia em DMEM suplementado com dexametasona 10-7 M (Sigma, St. Louis, MO, EUA), ácido ascórbico 50 μg / ml (Sigma, St. Louis, MO, EUA) e β-glicerol 10 mmol / l fosfato (Sigma, St. Louis, MO, EUA). A osteogênese foi avaliada por coloração com vermelho de alizarina.
O meio adipogênico consistia em DMEM suplementado com 0,5 mmol / l de 3-isobutil-1-metilxantina (IBMX) (Sigma, St. Louis, MO, EUA), 1 μmol / l de hidrocortisona (Sigma, St. Louis, MO, EUA), 0,1 mmol / l de indometacina (Sigma, St. Louis, MO, EUA). A diferenciação adipogênica foi avaliada por coloração com Oil Red O.
Identificação de antígenos de superfície hADSc por citometria de fluxo
Os hADSCs foram digeridos com tripsina e depois enxaguados duas vezes com DMEM, antes da ressuspensão a uma densidade celular de 2 × 10 7 células / ml. A suspensão de células (50 μl; 1 × 10 6 de 20 min a 37 ° C no escuro. O anti-CD34, anti-CD44 e anti-CD45 foram obtidos na CST (Beverly, MA, EUA); outros anticorpos foram obtidos da Abcam (Cambridge, MA, EUA). Em seguida, a suspensão de células foi centrifugada a × 500 g por 5 min, seguido pela remoção do sobrenadante e ressuspensão das células em 200 μl de tampão de coloração. Todas as etapas foram repetidas duas vezes antes da análise por citometria de fluxo.
Análise de imunoblotting (IB)
As células foram coletadas para imunotransferência conforme descrito anteriormente [14]. Os anticorpos primários usados foram anti-β-catenina (ab32572), anti-integrina β1 (ab30394) e anticolágeno II (ab34712), obtidos da Abcam (Cambridge, MA, EUA). Anti-β-actina (8H10D10, 1:2000), anti-GSK-3β (27C10, 1:1000) e anti-SOX (92G2, 1:1000) foram obtidos da Cell Signaling Technology (CST, Beverly, MA, EUA). Anticorpos conjugados com HRP secundários (1:1000–1:3000) foram adquiridos da CST.
Imunofluorescência
Para a diferenciação condrogênica, as células foram tratadas por 0, 6 e 12 dias, digeridas e cultivadas em vidro em placas de 24 poços (Costar353047, Corning, New York, EUA) por 24 h. As células foram lavadas duas vezes com solução tampão de fosfato gelada (PBS), fixada com paraformaldeído a 4% por 15 min em temperatura ambiente. Após o bloqueio, as células foram incubadas com o anticorpo primário reativo com integrina β1 por 1 h, seguido de incubação por 1 h no escuro com IgG anti-camundongo marcado com Alexa Fluor 488 (H + L) (CST # 4408, MA, EUA ), 4 ', 6-diamidino-2-fenilindole (DAPI, Sigma, MO, EUA). Para a coloração com faloidina, após o bloqueio, as células foram permeabilizadas com 0,2% de Triton X-100 por 30 min, em seguida, as células foram incubadas com DAPI e phalloidin-Alexa Farinha 573 (Life technologies, CA, EUA) por 1 h. Após três lavagens, a localização subcelular da integrina β1 e a alteração da actina filamentosa (F-actina) foram avaliadas durante a diferenciação da cartilagem com um microscópio confocal de varredura a laser (ZEISS, LSM 700, Oberkochen, Alemanha).
Preparação das dicas de AFM
As pontas de Si3N4 (DNP-10, Bruker Corp) com uma constante de mola (0,06 N / m) foram quimicamente modificadas pelo anticorpo anti-CD29 como segue [15]. As pontas foram limpas com acetona, luz ultravioleta e solução de piranha (H 2 SO 4 :H 2 O 2 =3:1, v / v ) para tempos diferentes (5 min, 30 min e 10 min). Após enxágue completo com água purificada, as pontas foram formadas por incubação com solução de 3-APTES 1% (Sigma, St. Louis, MO, EUA) em etanol por 30 min. As pontas foram lavadas três vezes com água ultrapura e tratadas com solução de glutaraldeído 2,5% (Sigma, St. Louis, MO, EUA) por 1 hora. O glutaraldeído supérfluo foi lavado três vezes com água. Finalmente, as pontas foram inseridas em uma solução de anti-integrina β1 (1 mg / ml) e incubadas durante a noite a 4 ° C. As sondas modificadas foram lavadas com PBS antes dos experimentos.
Medições AFM
AFM (Bioscope Catalyst, Bruker, EUA) foi usado para investigar a morfologia do hADSc e mudanças ultraestruturais durante a diferenciação condrogênica. A constante de força exata das pontas AFM foi medida em PBS. Para avaliar a morfologia e ultraestrutura, as células foram lavadas com PBS várias vezes. Em seguida, solução de paraformaldeído a 4% foi adicionada a um 2 de 3,5 cm prato de cultura por 15 min. Após as células serem lavadas com PBS, as células foram armazenadas em PBS a 4 ° C até serem utilizadas. A constante da mola das pontas variou de 4,2 a 5,8 N / m no modo de contato. Morfologia e imagens ultraestruturais de hADSc foram obtidas em PBS à temperatura ambiente por AFM. Imagens de ultraestrutura ao redor dos núcleos de hADSc foram obtidas no modo de contato. O software de análise de nanoscópio foi usado para avaliar a ultraestrutura da superfície celular para mais de 15 imagens 10 × 10 μm diferentes para pelo menos 15 células diferentes em (dia 0, 6, 12) grupos. A força de ligação entre as pontas AFM modificadas com integrina β1 e os receptores CD29 de hADSc vivo foi analisada durante diferentes períodos condrogênicos (0, 6 e 12 dias). A força de ligação foi medida no modo de aproximação-retração do sistema AFM (Bioscope Catalyst, Bruker, EUA). Para estudar os eventos de separação de células vivas com integrina β1, as pontas modificadas com anticorpo integrina β1 foram usadas em velocidades de retração de aproximação de 500 nm / s. A constante de força das pontas funcionalizadas foi de 0,058 ± 0,006 N / m. O limiar de força nas células foi de 800 pN. O anticorpo anti-integrina β1 (100 μg / ml) foi adicionado às células por 30 min antes dos experimentos de medição de força. O bloqueio da integrina β1 e as sondas nuas também foram utilizadas como controles para detectar a força de ruptura inespecífica entre as pontas e células modificadas com o anticorpo integrina β1. Para a quantificação da probabilidade de ligação do ligante − receptor da integrina β1, as curvas de força de interação específica foram medidas por sondas funcionalizadas com anticorpo integrina β1. Mais de 400 curvas de força foram medidas em um único experimento com resultados resumidos de pelo menos três experimentos independentes. Assim, aproximadamente 1200 curvas força-distância originais em cada experimento de comparação foram adquiridas de 30-40 células diferentes usando o software de análise Nanoscope do instrumento. Calculando a média dos valores de força para pelo menos três experimentos independentes, o efeito da indução condrogênica na força de interação entre o ligante integrina β1 e os receptores CD29 na superfície celular foi determinado.
Transcrição reversa e PCR em tempo real
Os kits de purificação de RNA TRIzol® Plus (Life Technologies, CA, EUA) foram usados e 1 μg de RNA foi transcrito reversamente para cDNA usando um kit de transcrição reversa de cDNA de alta capacidade (Invitrogen) de acordo com o protocolo do fabricante com pequenas modificações. Integrina β1 e GAPDH foram quantificados usando qRT-PCR com iniciadores específicos do gene:5′-TGGAGGAAATGGTGTTTGC-3 ′ (integrina β1-sentido) e 5′-CGTTGCTGGCTTCACAAGTA-3 ′ (integrina β1-antisense); 5′-CTGACTTCAACAGCGACACC-3 ′ (GAPDH-sense) e 5′-CCCTGTTGCTGTAGCCAAAT-3 ′ (GAPDH-antisense). Para PCR em tempo real, a PCR em tempo real Step One (Applied Biosystems) foi realizada usando Fast SYBR @ GREEN Master Mix (Life Technologies, CA, EUA). A expressão do gene alvo foi normalizada para GAPDH como um padrão interno e calculada usando o método comparativo 2-ΔΔCT. Cada ensaio foi conduzido em triplicado.
Análise estatística
Todos os experimentos foram realizados pelo menos três vezes, com dados expressos em média ± desvio padrão (DP). A comparação entre dois grupos foi conduzida por t teste. Diferenças significativas entre as médias dos grupos foram determinadas por análise ANOVA de uma via, seguida pelo teste T2 de Bonferroni e Tamhane (variâncias iguais não foram assumidas). Valores de p <0,05 foram considerados estatisticamente significativos.
Resultados e discussão
Avaliação de hADSc
As células-tronco mesenquimais são células estromais multipotentes com potencial osteogênico, adipogênico, condrogênico e miogênico. Existem dois meios principais para identificar hADSc, marcadores de CD de superfície celular e a capacidade de diferenciação [16]. Conforme mostrado no arquivo adicional 1:Figura S1 e arquivo adicional 2:Figura S2, as células derivadas foram hADSc. Em seguida, a proliferação celular da passagem 3 hADSc foi determinada pelo ensaio MTT (arquivo adicional 3:Figura S3).
Morfologia induzida e mudanças na ultraestrutura de superfície durante a condrogênese hADSc
AFM é sempre usado para detectar a morfologia celular e ultraestrutura em nanoescala [17]. A forma de uma célula está relacionada à sua função celular especializada e à organização do tecido. Em algumas pesquisas sobre câncer, o AFM pode ser usado como uma técnica de alta imagem para analisar mudanças morfológicas para a avaliação dos efeitos de drogas. Além disso, a forma das células-tronco mesenquimais é alterada durante a indução condrogênica [18]. Embora as mudanças na forma das células pareçam ser necessárias para a diferenciação, pouco se sabe sobre se a morfologia celular afeta os estágios iniciais de desenvolvimento da diferenciação das células-tronco mesenquimais. Portanto, alterações na morfologia e na ultraestrutura da membrana durante a condrogênese do hADSc foram avaliadas por AFM, uma vez que essas alterações são importantes [19] e podem influenciar diretamente a função das células [20]. A morfologia da superfície e a estrutura ultrafina de hADSc foram investigadas durante a diferenciação condrogênica por diferentes períodos de tempo (Fig. 1 e Fig. 2). A morfologia e a ultraestrutura da superfície eram obviamente diferentes em cada grupo de comparação. No dia 0, as células tinham uma forma de fuso alongada com uma superfície relativamente lisa. A arquitetura da membrana celular era homogênea. Após a indução condrogênica, nos dias 6 e 12, foram observadas alterações significativas na morfologia celular. A maioria das células encolheu gradualmente em uma forma poligonal (Fig. 1a) com uma diminuição na proporção média do comprimento / largura da célula durante a diferenciação condrogênica (Fig. 1b). Numerosos estudos mostram que as mudanças na morfologia celular são consistentes com o citoesqueleto das células [21]. Também encontramos alterações no citoesqueleto durante a diferenciação condrogênica, o que foi explicado nos últimos resultados.
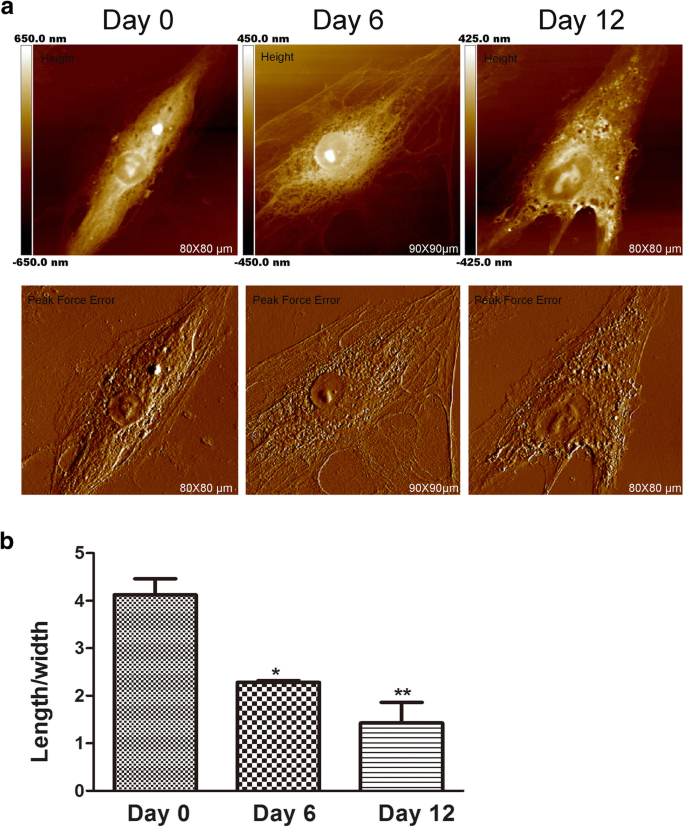
Características da morfologia do hADSc durante a condrogênese. a Imagens morfológicas de hADSc inteiro foram obtidas em 0, 6 e 12 dias de diferenciação condrogênica. As imagens foram analisadas por um modelo de imagem de erro de altura e força de pico da Nanoscope. b A relação comprimento / largura média das células foi medida após o tratamento de diferenciação condrogênica em 0, 6 e 12 dias. * p <0,05, ** p <0,01
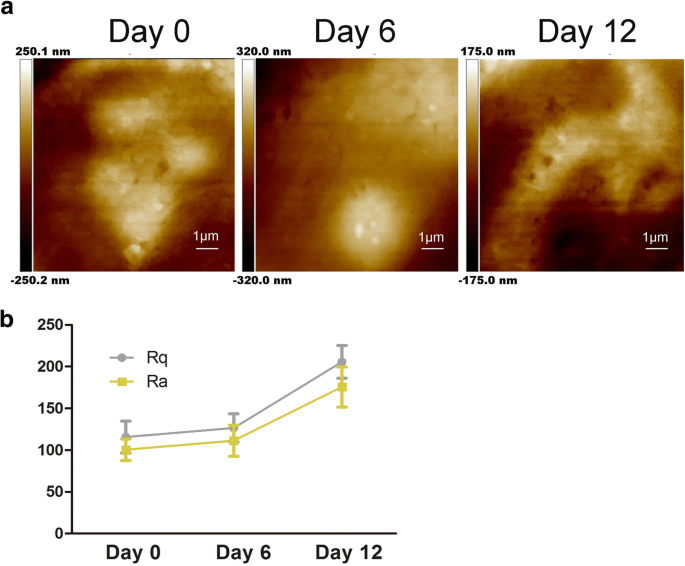
Características da ultraestrutura da membrana hADSc durante a diferenciação condrogênica. a Mudanças na ultraestrutura da membrana celular foram avaliadas após diferenciação condrogênica por 0, 6 e 12 dias. b Os parâmetros de rugosidade da superfície Ra e Rq das células foram medidos durante a indução condrogênica de hADSc por 0, 6 e 12 dias
Conforme mostrado na Fig. 2a, a ultraestrutura da membrana celular também mudou; as partículas tornaram-se aumentadas e heterogêneas. Estudos anteriores demonstraram que Ra e Rq foram os fabricantes do valor de rugosidade para avaliar a mudança em membranas celulares tratadas de maneiras diferentes [22]. Rq é sobre rugosidade quadrada média, \ (\ mathrm {Rq} =\ sqrt {\ frac {\ sum_ {t-1} ^ N {\ left (Zn- \ overline {Z} \ right)} ^ 2 } {N-1}} \); \ (\ mathrm {Rq} =\ sqrt {\ frac {\ sum _ {\ mathrm {t} -1} ^ {\ mathrm {N}} {\ left (\ mathrm {Zn} - \ overline {\ mathrm {Z }} \ right)} ^ 2} {\ mathrm {N} -1}}; \) Ra é sobre rugosidade média, \ (\ mathrm {Ra} =\ frac {1} {N} {\ sum} _ { t-1} ^ N1 \ mid Zi- \ overline {Z} \ mid \). Para obter a rugosidade, o tamanho da varredura é 10 μm × 10 μm. Como mostrado na Fig. 2b, tanto o Ra quanto o Rq de duas áreas diferentes aumentaram durante a condrogênese de hADSc. Os valores de Ra e Rq das células no dia 0 eram baixos, indicando uma superfície lisa (Fig. 2b). Os valores de Ra e Rq aumentaram concomitantemente com a diferenciação condrogênica, mostrando maior heterogeneidade e mais rugosidade na superfície celular (Fig. 2a). Com base nas mudanças observadas, a diferenciação condrogênica resultou em mudanças na morfologia celular e na relação altura / largura da célula (Fig. 1a, b). Existem estudos que mostram que a ECM pode regular a adesão celular regulando as integrinas [11]. Portanto, valores aumentados de rugosidade sugeriram mudanças na MEC e na ultraestrutura da membrana celular durante a condrogênese. Esses dados demonstram que a diferenciação condrogênica afeta a morfologia celular, a ECM e a estrutura da membrana celular.
Alterações do citoesqueleto durante a indução condrogênica de hADSc
Durante a diferenciação das células-tronco, a morfologia celular e as mudanças estruturais da membrana estão relacionadas ao citoesqueleto da célula, subsequente ao desenvolvimento de características celulares específicas da linhagem [21]. Como mostrado na Fig. 3a, os sinais de fluorescência vermelho e azul indicam respectivamente F-actina e DAPI. O citoesqueleto celular mudou muito durante a indução condrogênica na Fig. 3a. Por um lado, os microfilamentos do citoesqueleto percorreram o longo eixo da célula no grupo do dia 0, enquanto os microfilamentos do citoesqueleto se espalharam em uma matriz radial quando o hADSc foi tratado com diferenciação condrogênica por 12 dias. Por outro lado, a distribuição dos microfilamentos celulares foi homogênea no grupo do dia 0, mas os microfilamentos foram distribuídos principalmente na periferia do hADSc tratado com diferenciação condrogênica por 12 dias.
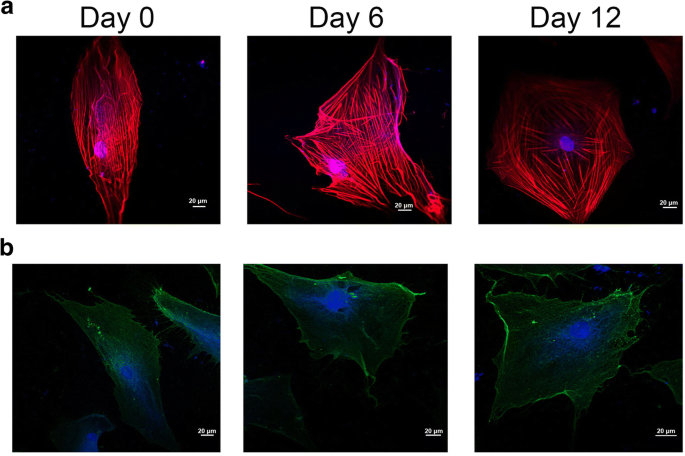
Organização do citoesqueleto e localização da integrina β1 na diferenciação condrogênica de hADSc. a Alterações no citoesqueleto foram detectadas durante a condrogênese de hADSc por microscopia confocal. b A localização da integrina β1 foi medida durante a diferenciação condrogênica por microscopia confocal. O citoesqueleto e o núcleo foram corados com F-actina e DAPI, respectivamente. Os sinais de fluorescência vermelho e azul indicam respectivamente F-actina e DAPI
Diferenciação condrogênica alterou a probabilidade de ligação da integrina β1 a receptores em hADSc
AFM também é uma ferramenta útil para o estudo da força de ligação entre ligantes e seus receptores, tornando clara a transdução do sinal do receptor de membrana na superfície das células [23]. Por AFM, as alterações entre a integrina β1 e seus receptores são medidas de forma visual, simples e específica. A interação do ligante β1 da integrina-receptor nas células vivas é uma forma de explorar o processo de ligação na membrana celular. O procedimento para a funcionalização da ponta de AFM é o acoplamento da integrina β1 às pontas de AFM por ligação de APTES e glutaraldeído. Estas pontas foram utilizadas para a detecção da ligação da integrina β1 aos receptores CD29 nas superfícies das células (Fig. 4a). A espectroscopia de força de molécula única (SMFS) foi usada para avaliar a distribuição da força de separação de células vivas anti-integrina β1 em regiões localizadas de hADSc vivo individual (Fig. 4b). As curvas de força representativas são mostradas na Fig. 4c, d, que representam uma curva de uma única molécula (Fig. 4c) e dois pares de curvas de pico de ruptura (Fig. 4d). Experimentos de bloqueio e experimentos com pontas de AFM nuas foram realizados para verificar a especificidade das curvas de força obtidas. As pontas nuas de AFM não detectaram nenhum pico de força específico (Fig. 4e). Experimentos simples de AFM mostraram que a probabilidade de ligação não específica da interação ligante β1 da integrina β1 na superfície do hADSc era inferior a 1%. Para os experimentos de bloqueio, o anticorpo anti-integrina β1 foi incubado com células por 30 min e, em seguida, as curvas de força foram registradas usando pontas funcionalizadas com integrina β1. O anticorpo bloqueador reduziu as curvas de força em 90% (Fig. 4f). Não houve diferença na probabilidade de ligação do receptor-ligante da integrina β1 nas superfícies das células entre os três grupos após o tratamento com anticorpo anti-integrina β1 (Fig. 4g). Estes resultados demonstram que as pontas de AFM modificadas com anticorpos foram muito úteis para detectar a força e que as pontas de AFM funcionalizadas com integrina β1 eram específicas.
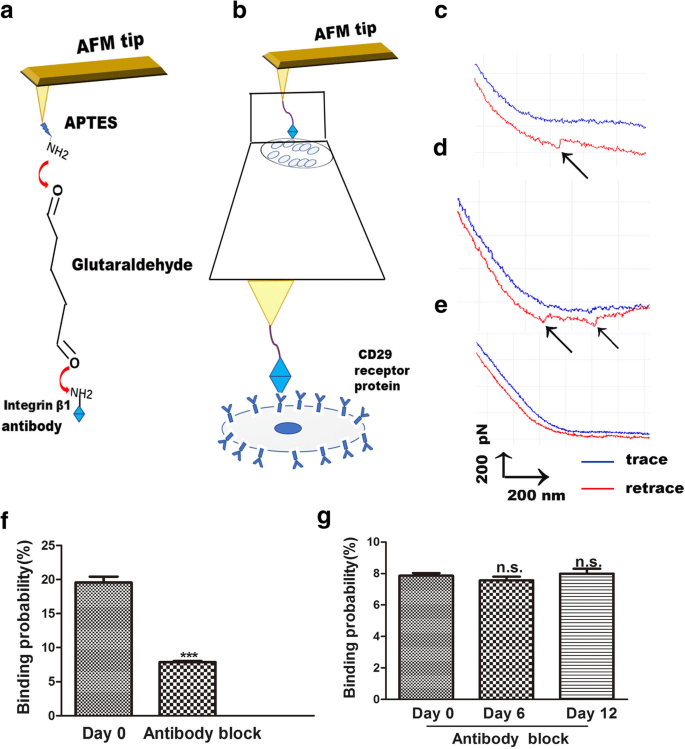
Medições de força AFM com ponta AFM funcionalizada com integrina β1 em hADSc vivo. a Representação esquemática da estratégia utilizada para imobilização da integrina β1 em uma ponta de AFM. b Representação esquemática da força de uma única molécula medida entre pontas AFM funcionalizadas com integrina β1 e hADSc vivo. c, d Curvas de força representativas obtidas com pontas AFM modificadas com integrina β1 em hADSc e e após o sistema ter sido bloqueado com a solução de anticorpo monoclonal integrina β1. f A probabilidade de ligação de pontas funcionalizadas com integrina β1 em hADSc antes e depois do bloqueio pelo anticorpo integrina β1 no dia 0. g A probabilidade de ligação de pontas funcionalizadas com CD29 em hADSc após o bloqueio pelo anticorpo integrina β1 em 0, 6 e 12 dias. *** p <0,001, n.s .. nenhuma diferença significativa
A força de ligação (força de ruptura) é a força de interação entre os ligantes e seus receptores [24]. Mudanças na morfologia e na ultraestrutura da superfície das membranas plasmáticas estão relacionadas a muitos processos da biologia celular, como diferenciação, apoptose e migração celular. Durante a diferenciação, acredita-se que as alterações no citoesqueleto estejam relacionadas às alterações da integrina, especialmente da integrina β1. A integrina β1 (CD29) é muito importante na adesão celular à ECM e na adesão célula-célula. Também pode interagir com proteínas intracelulares, estimulando moléculas sinalizadoras que estão relacionadas ao citoesqueleto de actina [25]. Neste estudo, alterações no citoesqueleto e na morfologia celular foram observadas durante a condrogênese hADSc por microscopia confocal de varredura a laser (CLSM) e AFM. Durante a diferenciação condrogênica, mudanças no citoesqueleto, morfologia e ultraestrutura da superfície podem ser um indicador novo e confiável do estado da célula. A integrina β1, o receptor CD29, é distribuída pela superfície celular conforme avaliado por imunofluorescência (Fig. 3b). A força de ligação e a estabilidade dos complexos ligante-receptor da integrina β1 durante a condrogênese hADSc foram avaliadas em 0, 6 e 12 dias de diferenciação. Um total de 1200 curvas foram registradas para cada dia, com forças de ruptura médias de 61,8 ± 22,2 pN, 60 ± 20,2 pN e 67,2 ± 22,0 pN, respectivamente (Fig. 5a – c). A distribuição da magnitude da força foi analisada como força média + SD (Fig. 5d). Não houve diferença significativa na força média entre os dias 0 e 6. Houve uma diferença na força média entre os dias 0 e 12. A magnitude da força de ligação aumentou no dia 12. Enquanto isso, eventos de ruptura em 0, 6 e 12 dias foram, respectivamente, 19,58 ± 1,74%, 28,03 ± 2,05% e 33,4 ± 1,89% (Fig. 5e). O aumento da probabilidade de ligação também indicou que a integrina β1 (CD29) desempenhou um papel importante na diferenciação condrogênica e pode fornecer informações para a diferenciação condrogênica, por meio de vias de sinalização. Portanto, nanodomínios de integrina β1 aumentados durante a diferenciação condrogênica podem afetar fundamentalmente a força de ligação do receptor-ligante CD29 em hADSc vivo. As mudanças na morfologia e na ultraestrutura da superfície das membranas plasmáticas acompanharam as mudanças na estrutura da proteína integrina β1, conformação, força de ligação e estabilidade dos complexos ligante-receptor da integrina β1 nas células. Em resumo, a integrina β1 desempenha um papel necessário na diferenciação condrogênica de hADSc.
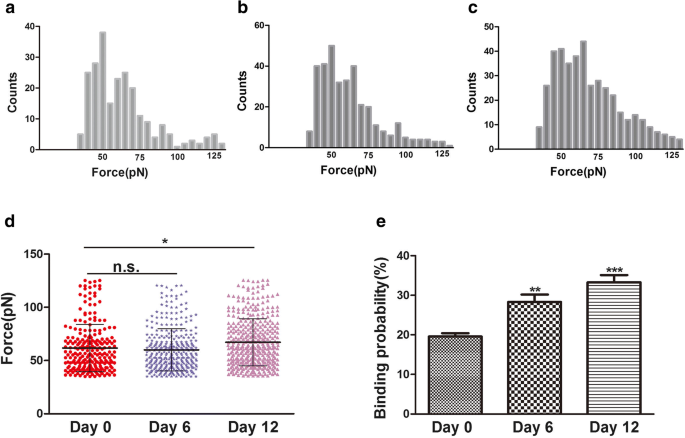
Força de ligação e probabilidade de ligação medidas na superfície de hADSc vivo por pontas AFM funcionalizadas com integrina β1. a – c Histogramas da força de ligação do receptor-anticorpo integrina β1 obtidos durante a diferenciação condrogênica de hADSc por 0, 6 e 12 dias. d As forças de ligação para os receptores β1 da integrina foram obtidas nos dias 0, 6 e 12 de diferenciação condrogênica de hADSc. e A probabilidade de ligação do receptor de integrina β1 foi detectada durante a diferenciação condrogênica de hADSc por 0, 6 e 12 dias. * p <0,05, ** p <0,01, *** p <0,001, n.s. nenhuma diferença significativa
Regulação positiva da integrina β1 durante a diferenciação condrogênica hADSc
Numerosos estudos demonstraram que os membros da família da integrina desempenham um papel importante na diferenciação celular. Além disso, as integrinas podem regular a interação entre o ambiente extracelular e as células, controlando as vias de transdução de sinal por meio de proteínas conectadas [26]. Estudos anteriores mostraram que a probabilidade de ligação pode ser afetada pela densidade e conformação da proteína transmembrana (receptores) na superfície celular [27]. A conformação da integrina pode ser uma antena fechada, que tem baixa afinidade para o ligante, ou uma antena aberta, que tem uma alta afinidade para o ligante [28, 29]. A expressão da integrina β1 foi regulada positivamente nos níveis transcricional e translacional com expressão aumentada do colágeno II, característica dos condrócitos (Fig. 6a, b). Como tal, a expressão da integrina β1 regulada positivamente foi consistente com o aumento da probabilidade de ligação, independentemente da conformação.

The role of integrin β1 and β-catenin/SOX pathway in regulating hADSc chondrogenic differentiation. a Protein integrin β1 was up-regulated during chondrogenesis of hADSc as assessed by western blotting. Cartilage differentiation up-regulated collagen II expression at different days. b The mRNA of integrin β1 was up-regulated during chondrogenic differentiation of hADSc. c Measurement of proteins associated with the β-catenin/SOX pathway during chondrogenic differentiation of hADSc for 0, 6, and 12 days. * p < 0.05, **p < 0.01
The Role of Integrin β1 in Chondrogenic Differentiation Regulated by the β-catenin/SOX Signaling Pathway
Previous studies have shown Wnt/β-catenin, PI3K, and mTOR signaling pathways to be related to integrin β1 [30,31,32]. Each is important in mesenchymal stem cell differentiation. Likewise, studies have demonstrated SOX and collagen II to be regulated by integrin β1 during chondrogenesis of hADSc. SOX is a hallmark component of the Wnt/β-catenin signaling pathway. Hence, we hypothesized that chondrogenic differentiation was regulated by the β-catenin/SOX pathway via integrin β1. SOX, GSK-3β, β-catenin, and integrin β1 were all increased during chondrogenesis of hADSc (Fig. 6c), with integrin β1 inducing cell signaling. These data demonstrate chondrogenic differentiation to be regulated by the β-catenin/SOX pathway via integrin β1.
Prospective and Limitations
In this work, changes in cellular morphology, the structure of the membrane, and the binding probability of integrin β1 ligand–receptors were demonstrated to be useful image markers to evaluate the chondrogenic differentiation process. This is a new method for evaluation of morphology, membrane ultrastructure, and changes in transmembrane proteins during chondrogenic differentiation. There are limitations to this study. Although increased binding probability was related to the high expression of integrin β1, the conformation of integrin β1 during chondrogenesis was not investigated. Further work is necessary to determine the conformation of integrin β1 during chondrogenic differentiation. Integrin β1 was demonstrated to participate in the β-catenin/SOX signaling pathway during chondrogenesis of hADSc. However, the relationship between integrin β1 and β-catenin/SOX signaling pathway is still not fully established. Further work is necessary to identify the exact role of integrin β1 in this pathway.
Conclusões
In the present work, a novel method (AFM) was employed to evaluate chondrogenic induction in hADSc. Cell surface ultrastructural changes were assessed by AFM imaging. AFM was used to investigate the binding force and binding probability between integrin β1 ligand and its receptors on the surface of hADSc by integrin β1-functionalized AFM tips. Based on AFM data, during chondrogenesis, cell morphology was changed from an elongated spindle shape to a polygonal shape with increased cell roughness. By use of integrin β1-functionalized AFM tips, the binding probability and force magnitude of integrin β1 ligand–receptor on the surface of hADSc were found to increase during chondrogenic induction. By immunoblot, integrin β1 was demonstrated to participate in the β-catenin/SOX signaling pathway, which regulated the chondrogenesis of hADSc. Taken together, these results and the established methodology contribute to a better understanding of cell morphology and roughness. Further, the data provide thermodynamic and kinetic insight into the integrin β1 ligand-binding process, at the single-molecule level. This AFM method will be useful for investigation of signaling pathways in living hADSc during chondrogenesis. Changes in the cellular nanostructure, as well as structure of the membrane, and the binding probability of transmembrane proteins are useful markers to evaluate chondrogenic differentiation mechanisms. This AFM method can be used to understand the mechanism of mesenchymal stem cell differentiation in tissue engineering and will be useful for an enhanced understanding of mesenchymal stem cell chondrogenic differentiation.
Abreviações
- AFM:
-
Força atômica microscópica
- CD:
-
Cluster of differentiation
- DAPI:
-
4′,6-diamidino-2-phenylindole
- DMEM:
-
Dulbecco’s Modified Eagle’s Medium
- ECM:
-
Extracellular matrix
- F-actin:
-
Filamentous actin
- FBS:
-
Soro fetal bovino
- hADSc:
-
Human adipose-deprived stem cells
- IB:
-
Immunoblotting analysis
- IBMX:
-
3-isobutyl-1-methylxanthine
- IGF-1:
-
Insulin-like growth factors-1
- ITS:
-
Insulin transferrin selenium
- mTOR:
-
Mammalian target of rapamycin
- OA:
-
Osteoartrite
- PBS:
-
Solução tampão de fosfato
- PI3K:
-
Phosphoinositide 3-kinase
- SD:
-
Desvio padrão
- SMFS:
-
Single-molecule force spectroscopy
- TGF-β1:
-
Transforming growth factor-beta1
Influência da largura do poço quântico nas propriedades de eletroluminescência de diodos emissores de luz ultravioleta profunda de AlGaN em diferentes temperaturas
Uma nova microestrutura de cluster de nanocônios com propriedades anti-reflexo e superhidrofóbicas para dispositivos fotovoltaicos
Nanomateriais
- Demonstração de um biossensor flexível à base de grafeno para detecção sensível e rápida de células cancerosas do ovário
- Tecnologias de Deposição de Camada Atômica Avançada para Micro-LEDs e VCSELs
- Nanopartículas de fósforo preto promovem a diferenciação osteogênica de EMSCs por meio da expressão de TG2 regulada positivamente
- Revelando a estrutura atômica e eletrônica das nanofibras de carbono de copo empilhado
- Análise de Actina e Organização de Adesão Focal em Células U2OS em Nanoestruturas Poliméricas
- Nanotécnicas Inativam células-tronco cancerosas
- Comparação de eletrodos metálicos padronizados do tipo nanofuro e nanopilar incorporados em células solares orgânicas
- Síntese de nanocristais de ZnO e aplicação em células solares de polímero invertido
- Estudo da Força de Adesão e Transição de Vidro de Filmes de Poliestireno Fino por Microscopia de Força Atômica
- Uma Sonda Fluorescente Resumível BHN-Fe3O4 @ SiO2 Híbrida Nanoestrutura para Fe3 + e sua Aplicação em Bioimagem