


Avaliação da atividade do citocromo P450 3A4 inibido por nanopartículas de ouro e dos mecanismos moleculares subjacentes à sua toxicidade celular na linha celular C3A do carcinoma hepatocelular humano
Resumo
As interações das nanopartículas de ouro de 40 e 80 nm (AuNP) funcionalizadas com polietilenimina catiônica ramificada (BPEI), ácido lipóico aniônico (LA) ou polietilenoglicol neutro (PEG) com a linha celular C3A de carcinoma hepatocelular humano (HCC) foram investigadas no ausência e presença de corona de proteína plasmática humana (PC). Todos os AuNP nus (sem PC) além de LA-AuNP de 80 nm foram citotóxicos para C3A, mas PC atenuou suas citotoxicidades. A captação celular dependente do tempo de AuNP aumentou além de BPEI-AuNP de 40 nm, mas o PC suprimiu sua captação além de PEG-AuNP de 80 nm. Respostas bifásicas de estresse oxidativo / nitrosativo por BPEI-AuNP ocorreram em células C3A, enquanto PEG-AuNP foi um potente antioxidante. Todo o AuNP puro inibiu a atividade do citocromo P450 (CYP) 3A4, independentemente do tamanho e da carga superficial, mas o PC recuperou sua atividade além do PEG-AuNP. A expressão do gene modulado por PEG-AuNP de 40 nm esteve principalmente envolvida na β-oxidação de ácidos graxos mitocondriais e em um menor grau de transportadores de efluxo / captação hepática. Esses estudos contribuem para uma melhor compreensão da interação do AuNP com os principais processos biológicos e seus mecanismos moleculares subjacentes no CHC, que podem estar ainda mais implicados no desenvolvimento de um alvo terapêutico mais eficaz no tratamento do CHC.
Histórico
O carcinoma hepatocelular (CHC) é um dos cânceres mais comuns em todo o mundo e a causa de mortalidade por câncer que mais cresce nos EUA [1, 2]. Dado que o CHC foi diagnosticado em estágios avançados, os tratamentos curativos do CHC incluem transplante de fígado ou ressecção cirúrgica no início do desenvolvimento do tumor e quimioterapia e radioterapia para um estado avançado do tumor. O HCC freqüentemente desenvolve uma alta resistência aos agentes antineoplásicos convencionais, uma molécula citotóxica não seletiva que pode resultar em efeitos adversos sistêmicos. Os avanços recentes em terapia gênica, ou seja, terapia gênica baseada em RNA de interferência (RNAi), têm sido utilizados no atual tratamento de CHC [3, 4]. A eficácia do RNAi requer que o vetor seja entregue ao interior da célula-alvo [5]. Os vetores para uma entrega de gene com sucesso são vetores virais e não virais. Os vírus oferecem maior eficiência de entrega de genes, mas os vetores não virais são preferidos devido a questões de segurança com os vetores virais. Nanopartículas (NP) como vetores não virais para entrega de genes direcionados ou sistema de entrega de drogas ganharam grande atenção para melhorar a eficiência terapêutica e diminuir a toxicidade nos níveis sistêmicos e / ou celulares no tratamento de HCC [4, 6]. Assim, torna-se muito importante identificar o mecanismo molecular e a via biológica subjacente ao distúrbio celular e à toxicidade do NP em células e tecidos alvo. Estudos recentes in vitro demonstraram que o perfil de expressão gênica combinado com respostas celulares e bioquímicas forneceu uma avaliação direta da perturbação celular e potencial toxicidade NP [7,8,9,10].
As nanopartículas de ouro (AuNP) têm sido usadas como veículo de entrega para entrega específica de alvos de frações de silenciamento de genes, sozinhas ou em combinação com outras drogas devido às suas propriedades físico-químicas únicas e químicas de superfície [11, 12]. A interação do AuNP com as proteínas do plasma sanguíneo forma a coroa protéica, que por sua vez altera a química da superfície do NP e influencia as respostas biológicas subsequentes, como a absorção celular e potencial toxicidade [13, 14]. A absorção celular de AuNP em diferentes linhagens de células cancerígenas humanas e células primárias foi criticamente afetada pela formação da corona da proteína, independentemente do tamanho e da carga superficial [7,8,9, 14,15,16,17].
O estresse oxidativo dependente do tamanho e da carga superficial também foi observado na linha de células de câncer de mama humano, MDA-MB-231, carcinoma hepatocelular HepG2 e células de leucemia humana HL-60 em resposta a AuNP, que foram associadas à citotoxicidade de NP [18 , 19]. A citotoxicidade induzida por AuNP ocorreu em várias linhagens de células de câncer humano e células humanas primárias de uma maneira específica para o tipo de célula [7,8,9, 20, 21].
As enzimas do citocromo P450 (CYP) desempenham um papel importante na bioativação ou inativação de numerosos fármacos citotóxicos e na susceptibilidade do hospedeiro à carcinogenicidade de fármacos anticancerígenos [22]. AuNP influenciou a atividade catalítica das enzimas CYP nos níveis celular e molecular in vivo e in vitro [7, 23,24,25]. AuNP exibiu consideravelmente a expressão gênica diferencial predominantemente envolvida em marcadores de estresse oxidativo na linha celular de fibroblastos de pulmão humano MRC-5 e disfunção mitocondrial em células da veia umbilical humana (HUVEC) e hepatócitos humanos, que se correlaciona com um aumento na produção de peróxido de lipídio e um alta citotoxicidade [8, 9, 26]. Embora este conhecimento sugira reciprocamente que AuNP causa morte celular apoptótica ou necrótica em vários tipos de células e altera as funções celulares e bioquímicas combinadas com a expressão gênica diferencial em vias de resposta ao estresse e toxicidade, as vias específicas através das quais AuNP exerce seus efeitos tóxicos dentro da célula ou biológicos sistema permanecem desconhecidos.
Aqui, este estudo investigou os efeitos da coroa da proteína, tamanho e carga superficial na interação do AuNP com a célula HCC humana C3A. Primeiramente, a captação celular dependente do tempo de 40 e 80 nm AuNP funcionalizada com BPEI catiônico, ácido lipóico aniônico (LA) ou polietilenoglicol neutro (PEG) em células C3A foi determinada com e sem corona de proteína plasmática humana (PC). Em segundo lugar, a citotoxicidade induzida por AuNP e a produção de espécies reativas de oxigênio (ROS) / espécies reativas de nitrogênio (RNS) foram monitoradas juntamente com seus efeitos inibitórios na atividade de CYP3A4. Por último, o mecanismo de ação molecular associado à toxicidade AuNP foi caracterizado usando o Human Molecular Toxicology Pathway Finder e o Human Drug Transporters RT 2 Matriz de PCR Profiler ™.
Métodos
Síntese de nanopartículas de ouro
O BPEI catiônico de 40 e 80 nm, LA aniônico e PEG Biopure ™ AuNP neutro foi sintetizado de forma personalizada a partir de nanoComposix (San Diego, CA). Tamanho de partícula, índice de polidispersidade (PDI) e zeta (z) -potencial e propriedades espectrais foram caracterizados com espalhamento de luz dinâmico (DLS), microscopia eletrônica de transmissão (TEM) e espectroscopia UV-Vis. AuNP foi sintetizado através da redução do hidrato de tetracloroaurato (III) de hidrogênio em solução aquosa de carbonato de potássio seguido do processo de envelhecimento e filtração tangencial (TFF). A superfície AuNP foi funcionalizada com LA ou PEG pela adição de ácido di-hidrolipóico (0,2:1, w / w ) ou PEG terminado com tiol-metoxi (Laysan Bio Inc., Arab, AL) (0,5:1, w / w ), respectivamente, seguido por lavagem TFF e filtração estéril. Superfícies funcionalizadas com BPEI de AuNP foram sintetizadas via química de EDC ligando o ácido carboxílico de LA a aminas livres de BPEI seguido por lavagem de TFF e subsequente centrifugação para uma remoção de BPEI não ligado.
Preparação da proteína corona
Plasma sanguíneo humano agrupado ( n =5) foram obtidos na Biological Speciality Corp. (Colmar, PA). AuNP foram incubados em plasma humano (55%, v / v ) a uma velocidade constante de 250 rpm a 37 ° C por 1 h, conforme relatado [7, 8]. As proteínas não ligadas e fracamente associadas foram removidas por lavagens repetidas com solução salina tamponada com fosfato (PBS) a 20.000 × g por 20 min a 20 ° C. O AuNP revestido com corona (PC) de proteína plasmática final humana foi disperso em PBS e, em seguida, diluído em meio de cultura de células para posterior caracterização físico-química ou dosagem. O protocolo detalhado é fornecido no arquivo Adicional 1.
Caracterização físico-química de AuNP
Diâmetros hidrodinâmicos ( D H ), PDI e potencial z de 40 e 80 nm nu (sem PC) AuNP funcionalizado com BPEI, LA e PEG em água desionizada (DI) foram analisados a 25 ° C em 0 h usando o Zetasizer Nano ZS (Malvern Instruments, Worcestershire, UK); para AuNP revestido com PC em PBS a 25 ° C a 0 h; e para todos os AuNP nus e PC em meio de cultura de células completo a 37 ° C em 0 he 24 h. O meio de cultura de células completo continha meio essencial mínimo de Eagle (EMEM) suplementado com 10% de FBS (ATCC ® , Manassas, VA). Uma amostra foi medida 5 vezes com 11 subexecuções de 10 s cada. Além disso, os espectros de absorção óptica foram medidos usando o leitor de microplaca híbrido multimodo Synergy H1 (BioTek Instruments Inc., Winooski, VT) em temperatura ambiente em 0 h.
Microscópio Eletrônico de Transmissão
A morfologia AuNP foi caracterizada usando TEM. Toda a solução nua e PC AuNP (5 μL) foi colocada em grades de cobre de malha 200 seguida por secagem ao ar em temperatura ambiente. As amostras foram visualizadas em um Tecnai G2 Spirit BioTWIN com um detector Oxford (FEI Company, Hillsboro, OR) a uma tensão de aceleração de 120 kV. O conjunto de microscopia GATAN (GATAN Inc., Pleasanton, CA) mediu os diâmetros AuNP.
Cultura de células e medição de viabilidade
Células C3A de carcinoma hepatocelular humano (ATCC ® CRL-10741 ™) foram adquiridos da ATCC ® (Manassas, VA), cultivado em EMEM completo (ATCC ® , Manassas, VA) suplementado com FBS a 10% e expandido para confluência de aproximadamente 80% em frasco T75 com mudanças de meio a cada 4 dias. Depois de 0,25% ( w / v ) tripsina-digestão com ácido etilenodiaminotetracético (EDTA) 0,53 mM, as células foram plaqueadas em placas de 96 poços em 8 × 10 4 células por poço e incubadas a 37 ° C em uma atmosfera umidificada de 95% de ar e 5% de CO 2. Após 48 h de incubação, as células foram dosadas com AuNP na ausência e presença de PC. As células C3A entre as passagens 9 e 12 foram utilizadas para a dosagem.
A viabilidade C3A foi determinada usando o alamarBlue ® ensaio de viabilidade (Thermo Sci., Waltham, MA) conforme descrito [7, 27]. As células nas placas de 96 poços foram tratadas com BPEI-, LA- e PEG-AuNP de 40 e 80 nm com e sem PC variando de 0 a 250 μg / cm 2 . Após 24 h, 10% de alamarBlue ® reagente em EMEM completo ( v / v ) foi adicionado à cultura de células e incubado durante 3 h a 37 ° C. O EMEM completo serviu como dispersante. As interações de AuNP com o ingrediente ativo de alamarBlue ® reagente, resazurina ou um produto reduzido, resorufina foram medidos como controles. AuNP e resazurina (sem células) ou meio de manutenção (sem células) serviram como controles de fundo. A fluorescência, proporcional à viabilidade celular, foi normalizada para controles e expressa como uma porcentagem em relação ao grupo de células de controle.
Medição de captação celular com espectrometria de massa de plasma indutivamente acoplado
As células foram semeadas em 8 × 10 4 células por poço de placas de 96 poços e dosadas com concentração não tóxica de 1,56 μg / cm 2 de todos os nus e PC AuNP por 0,5, 1, 3, 6, 12 e 24 h. A etapa de ataque foi incorporada para remover AuNP ligado à membrana celular e sua ligação não específica aos poços, como relatado anteriormente [28]. A colheita de células foi seca e digerida em água régia e a concentração de Au intracelular foi quantificada usando o NexION ™ Espectrometria de massa com plasma acoplado indutivamente 350X (ICP-MS) (PerkinElmer, Waltham, MA). A captação celular de AuNP foi calculada conforme relatado anteriormente e expressa como o número de AuNP por célula [29]. O protocolo detalhado é fornecido no arquivo Adicional 1.
Medições de tensão oxidativa / nitrosativa
As células foram semeadas em 8 × 10 4 células por poço de placas de 96 poços e dosadas com BPEI- e PEG-AuNP de 40 nm até 125 μg / cm 2 por 1, 3 e 24 h. A medição direta do estresse de oxigênio / nitrosação foi testada com o kit de ensaio de espécies reativas de oxigênio total (ROS) / superóxido (SO) (Enzo Life Sciences, Farmingdale, NY) como descrito anteriormente [30]. Fluorescência, proporcional ao aumento em ROS / espécies reativas de nitrogênio (RNS) (Ex488 / Em520 nm) ou SO (Ex550 / Em610 nm) foram medidas com o leitor de microplaca. O protocolo detalhado é fornecido no arquivo Adicional 1.
Atividade do citocromo P450 3A4
Os efeitos adversos de 40 e 80 nm nu e PC AuNP na atividade de CYP3A4 foram caracterizados usando ensaios P450-Glo ™ (Promega Corp., Madison, WI) como totalmente descrito [7]. As células C3A em placas de 96 poços foram dosadas na concentração letal média (LC 50 ) valores:127,3 μg / cm 2 do BPEI-AuNP de 40 nm, 205,5 μg / cm 2 do BPEI-AuNP de 80 nm, 192,5 μg / cm 2 do LA-AuNP de 40 nm e 129,5 μg / cm 2 do PEG-AuNP de 40 nm. Desde LC 50 valores de 80 nm LA- e PEG-AuNP não foram determinados, as células foram tratadas com LC 50 valores de 40 nm LA- e PEG-AuNP (192,5 μg / cm 2 e 129,5 μg / cm 2 , respectivamente). Após o final da incubação de 24 h, as células foram incubadas com um substrato CYP3A4 (luciferina-IPA) a 37 ° C por 3 h. O sinal de luminescência, proporcional a uma atividade enzimática, foi medido com um leitor de microplaca e então normalizado para controles. Os controles foram atribuídos para avaliar a interação de AuNP com substratos ou metabólitos e substratos livres de células. A atividade de CYP foi expressa como uma porcentagem em relação ao grupo de células de controle.
Perfil de expressão genética
Uma vez que PEG-AuNP de 40 nm tóxico foram empregados na inibição da atividade do CYP3A4 e da atividade antioxidante em células C3A com alta captação celular, ele foi selecionado para caracterizar os mecanismos moleculares de ação subjacentes à sua toxicidade e respostas celulares diferenciais. As células foram semeadas em 2,5 × 10 6 células por poço de placas de 6 poços e dosadas com LC 50 valor do PEG-AuNP de 40 nm durante 24 h a 37 ° C. No final da incubação, as células foram submetidas ao isolamento de RNA e, em seguida, a síntese de cDNA usando RNA total com um valor médio de números de integridade de RNA (RIN) de 7,8 foi conduzida conforme descrito anteriormente [7,8,9]. O cDNA resultante foi misturado com RT 2 SYBR green master mix (Qiagen Inc., Valencia, CA) e depois aplicada ao Human Molecular Toxicology Pathway Finder ou Human Drug Transporters RT 2 Matrizes de PCR Profiler ™ em Quantstudio ™ 7 Flex (Applied BioSystem, Foster City, CA). Genes expressos diferencialmente com a mudança de dobra <- 2 e> 2 e um p <0,05 representou a regulação para baixo e para cima do gene de interesse. Para validar o RT 2 Para dados da matriz de PCR, uma expressão de nove genes selecionados foi avaliada com a síntese de cDNA e subsequente PCR em tempo real. As sequências de primer estão resumidas no arquivo adicional 1:Tabela S1. Todas as reações de PCR foram realizadas em triplicado. O protocolo detalhado de condições de PCR em tempo real e quantificação é fornecido no arquivo adicional 1.
Análise estatística
Concentração letal média (LC 50 ) os valores de AuNP em células C3A foram estimados ajustando uma equação de Hill com inclinação variável aos dados observados (a entrada dos níveis de concentração de AuNP e a viabilidade celular correspondente) usando GraphPad Prism 6 (La Jolla, CA), conforme descrito [7]. A análise de variância unilateral (ANOVA) foi conduzida usando SAS 9.4 (SAS Institute, Cary, NC) para avaliar os efeitos de AuNP na produção de ROS / RNS e captação celular em células C3A. Se significativo, a comparação múltipla foi realizada com o teste de diferença significativa honesta (HSD) de Tukey em um p <0,05.
Resultados e discussão
Caracterização físico-química de PC AuNP de plasma nu e humano
Os efeitos do tamanho de NP, carga de superfície e formação de PC de plasma humano em torno de AuNP no diâmetro hidrodinâmico (D H ), índice de polidispersidade (PDI), potencial z e uma propriedade espectral, bem como a morfologia, foram caracterizados usando espectroscopia DLS, TEM e UV-Vis (Fig. 1). Em imagens TEM, todo o AuNP puro (sem PC) em água DI foi monodisperso com a distribuição de tamanho apertada e faixas de espectro UV-Vis exclusivas de 521–553 nm (Fig. 1a). As formações de PC em torno de AuNP foram observadas com as mudanças na distribuição de tamanho e redshifts dos espectros de absorção (Fig. 1b). O D H e os valores de PDI de 40 e 80 nm nu e PC AuNP em EMEM completo foram compatíveis até 24 h a 37 ° C, exceto para PC BPEI-AuNP de 40 e 80 nm que mostraram uma diminuição nos valores de PDI (0,29 e 0,32, respectivamente ) a 24 h a 37 ° C em comparação com aqueles a 0 h a 37 ° C (0,62 e 1,0, respectivamente) (Tabela 1). Os valores de potencial Z de todos os AuNP nus e PC diminuíram relativamente em 24 h a 37 ° C em comparação com aqueles em 0 h a 37 ° C. Uma agregação de 40 e 80 nm PC BPEI-AuNP em PBS e EMEM completo foi observada, que se correlacionou com múltiplos picos em uma distribuição de tamanho e mudanças em D H e redshifts do espectro de absorbância em relação ao BPEI-AuNP nu (Fig. 1 e arquivo adicional 1:Figura S1, Tabela 1). Esses resultados foram apoiados por estudos recentes de que o PC de 40 e 80 nm e o BPEI-AuNP revestido por corona com albumina de soro humano foram agregados em PBS e vários meios de cultura de células [7,8,9].
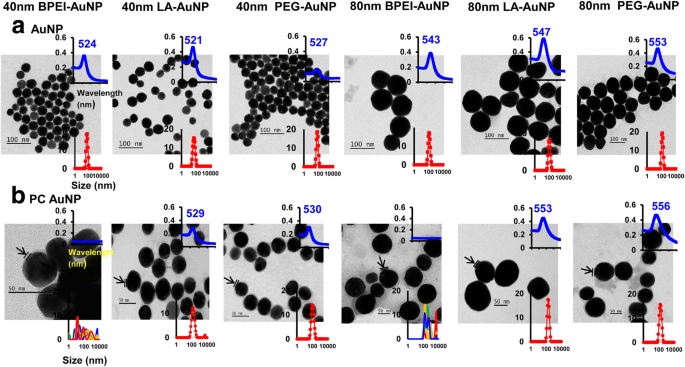
Micrografias eletrônicas de transmissão de a AuNP em água deionizada e b PC AuNP em PBS a 0 h a 25 ° C, comprimento de onda do espectro de UV-Vis (inserção superior) e a distribuição de espalhamento de luz dinâmica (inserção inferior). As setas indicam a formação do PC. PC corona de proteína plasmática humana, BPEI polietilenimina ramificada, LA ácido lipóico, PEG polietileno glicol
Citotoxicidade AuNP
A citotoxicidade de AuNP foi medida usando a concentração letal média (LC 50 ) em células C3A. A carga de superfície NP-, tamanho de partícula- e formação de PC em torno de LC dependente de NP 50 análises com AuNP foram mostradas na Fig. 2. Todos os 40 nm BPEI-, LA e PEG-AuNP e 80 nm BPEI-AuNP foram citotóxicos para células C3A com o LC 50 correspondente varia de 127,3 a 205,5 μg / cm 2 (Fig. 2a). O PEG-AuNP nu de 80 nm exibiu 59% de viabilidade celular na concentração mais alta de 250 μg / cm 2 , enquanto o LA-AuNP de 80 nm não foi citotóxico. PC reduziu a toxicidade de AuNP em função do tamanho e modificação de carga superficial, exceto para BPEI-AuNP de 80 nm que mostrou 51% de viabilidade celular a 250 μg / cm 2 às 24 h (Fig. 2b). Estudos recentes demonstraram que o BPEI-AuNP nu de 40 nm foram tóxicos para hepatócitos humanos primários, HUVEC e células tubulares proximais renais humanas (HPTC) (LC 50 varia de 22,4–80,3 μg / cm 2 ) [7,8,9]. O BPEI-AuNP revestido com PC foi citotóxico para hepatócitos humanos, mas AuNP revestido com HSA não foi citotóxico [7]. Esses resultados sugeriram que as células C3A são mais resistentes à toxicidade de AuNP do que as células humanas primárias devido a uma alta taxa de proliferação e atividade metabólica da linha celular cancerosa [31].
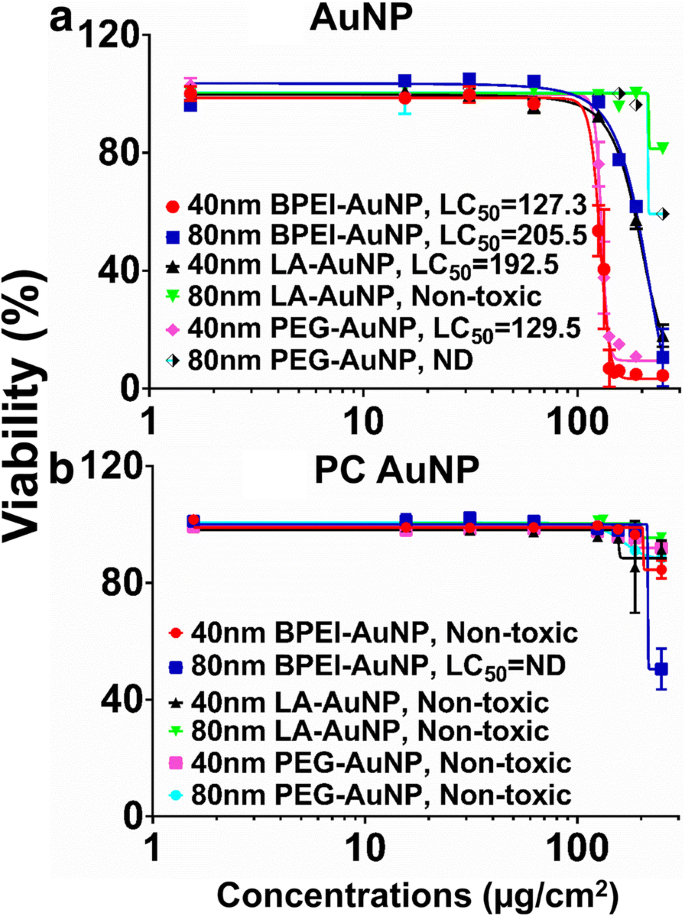
Viabilidade C3A e LC 50 valores de 40 e 80 nm a AuNP e b PC AuNP. Os dados representam a média ± D.P. ( n =3). Corona de proteína plasmática humana PC, ND não determinado, polietilenimina ramificada BPEI, ácido LA lipóico, polietilenoglicol PEG, LC 50 concentração letal mediana
Captação intracelular de AuNP
A absorção celular dependente do tamanho de NP, carga superficial e PC de todos os AuNP nus e PC foram determinados em 1,56 μg / cm 2 até 24 h. ANOVA mostrou mudanças significativas com o tamanho, PC e tempo ( p <0,0001) e interações (PC × tamanho, PC × tempo, tamanho × tempo e PC × tamanho × tempo) ( p <0,001) para toda a captação de AuNP, além de uma interação insignificante (PC × tamanho) para a captação de LA e PEG-AuNP ( p =0,2). Como mostrado na Fig. 3a-f, um aumento linear na captação celular de 40 e 80 nm nu e PC AuNP foi observado além de 40 nm nu e PC BPEI-AuNP que atingiu a maior captação celular em 6 h e diminuiu depois ( Fig. 3a). No entanto, em 24 h, o BPEI-AuNP catiônico de 40 nm continha a maior captação seguido por PEG-AuNP de 40 nm neutro e LA-AuNP de 40 nm aniônico, que foi associado com a ordem de citotoxicidade de células C3A de AuNP (Fig. 2a ) Este resultado é consistente com os estudos anteriores de que poli (N- (2-aminoetil) acrilamida) catiônico e BPEI-AuNP tiveram a maior captação celular em comparação com aqueles de poli (ácido acrílico) aniônico - e LA-AuNP e poli neutro ( N- (2,3-dihidroxipropil) acrilamida e PEG-AuNP em células Caco-2 de adenocarcinoma colorretal humano, HPTC e hepatócitos humanos [9, 32]. Além disso, o complexo NP-PC atenuou todos os BPEI- de 40 e 80 nm e LA-AuNP e o PEG-AuNP de 40 nm em células C3A, mas acelerou a captação de PEG-AuNP de 80 nm (Fig. 3f). Estes resultados são suportados pelos estudos recentes que o PC inibiu a captação de AuNP em HUVEC, HEK e HPTC, independentemente do tamanho e da carga superficial [8, 9, 33]. Em contraste, PC e HSA coronas aumentaram a captação de PEG-AuNP de 40 nm em hepatócitos humanos, mas este último induziu a captação de PEG-AuNP de 80 nm em HEK [7, 33] .
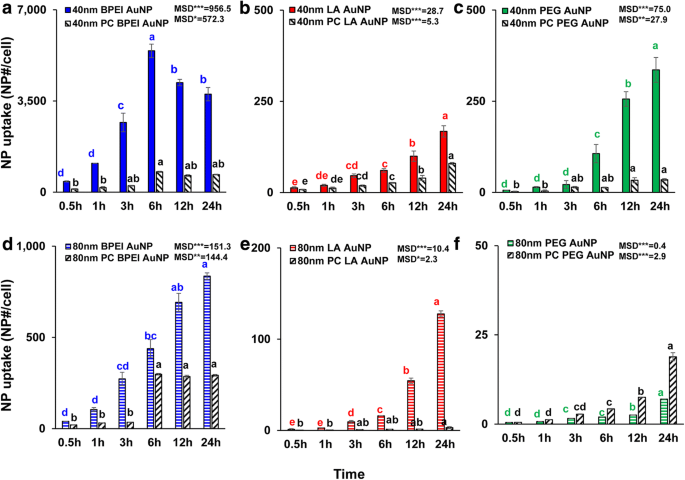
Captação celular dependente do tempo de 40 nm a BPEI-AuNP, b LA-AuNP e c PEG-AuNP, e 80 nm d BPEI-AuNP, e LA-AuNP e f PEG-AuNP na ausência e presença de PC em células C3A por até 24 h. Os dados representam a média ± D.P. ( n =3). As letras eram significativamente diferentes de acordo com o teste HSD de Tukey. BPEI polietilenimina ramificada, LA ácido lipóico, PEG polietilenoglicol, PC corona de proteína plasmática humana, MSD diferença mínima significativa. * p <0,05; ** p <0,005; *** p <0,0001
Medições de estresse oxidativo e nitrosativo
Uma vez que o BPEI- e PEG-AuNP nu de 40 nm exibiram a maior citotoxicidade e captação celular em células C3A em comparação com outro AuNP, eles foram selecionados para investigar o estresse oxidativo / nitrosativo induzido por AuNP. Ambos AuNP modulou a geração de ROS / RNS em células C3A de maneira dependente do tempo e da concentração ( p <0,0001) e por interação (tempo × concentração, p <0,0001). Como mostrado na Fig. 4a, a geração de ROS / RNS diminuiu nas concentrações mais altas de 40 nm BPEI-AuNP (62,5 μg / cm 2 e 125 μg / cm 2 ) em 1 h a 37 ° C, mas aumentou até 24 h. Em contraste, o PEG-AuNP de 40 nm suprimiu substancialmente a geração de ROS / RNS a 39,1 μg / cm 2 em diante com uma mudança de dobra <0,5 até 24 h (Fig. 4b). A ativação da morte celular freqüentemente contribui para a toxicidade de NP e, na maioria dos casos, um aumento na produção de ROS / RNS, levando ao estresse oxidativo, é responsável pela toxicidade de NP [34]. A produção de ROS / RNS dependente da carga superficial foi observada com BPEI catiônico de 40 nm e PEG-AuNP neutro. O BPEI-AuNP de 40 nm mostrou um padrão bifásico de geração de ROS / RNS (antioxidante em 1 h e pró-oxidante em 3 h em diante) em altas concentrações, que foi associado à sua citotoxicidade em células C3A (Fig. 2a). Este resultado é consistente com os estudos anteriores de que a geração de ROS induzida por BPEI-AuNP de 40 e 80 nm e citrato-AuNP de 20 nm foi associada com suas citotoxicidades em hepatócitos humanos e células HepG2, respectivamente, de maneira dependente do tempo e da concentração [7, 35]. AuNP mostrou citotoxicidade induzida por estresse oxidativo em células humanas de leucemia promielocítica, HL-60 com redução total da glutationa, independentemente do tamanho [19]. Em contraste, o PEG-AuNP de 40 nm serviu como um antioxidante sugerindo que o estresse oxidativo / nitrosativo pode não ser um mecanismo direto da citotoxicidade induzida por PEG-AuNP de 40 nm em células C3A (Fig. 2b).
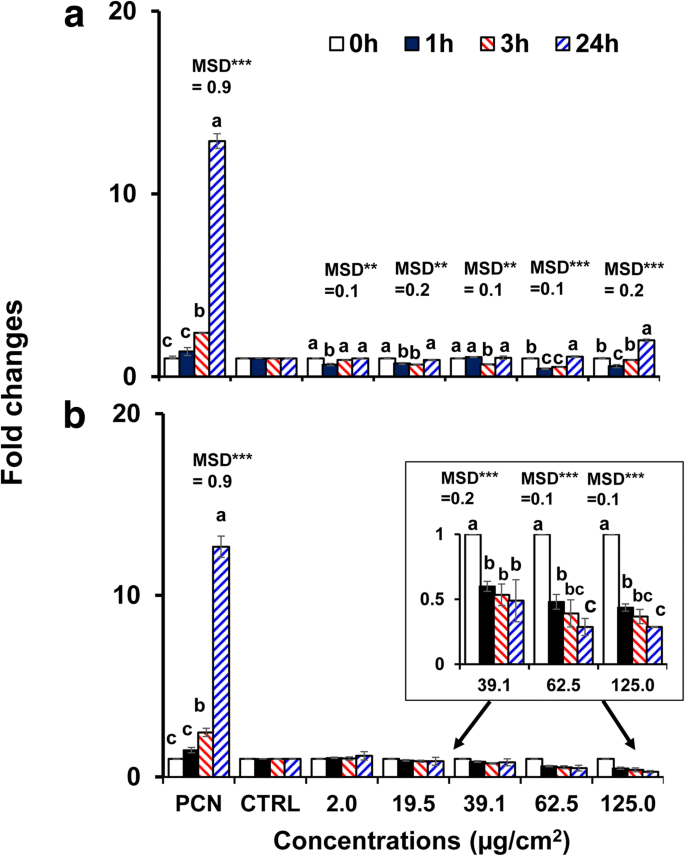
Produção de ROS / RNS dependente do tempo e da concentração em células C3A expostas a a o BPEI-AuNP de 40 nm e b o PEG-AuNP de 40 nm até 24 h. Os dados representam a média ± D.P. ( n =3). As letras eram significativamente diferentes de acordo com o teste HSD de Tukey. BPEI polietilenimina ramificada, LA ácido lipóico, PEG polietilenoglicol, CTRL controle, MSD uma diferença mínima significativa, PCN piocianina (indutor de ROS). ** p <0,005; *** p <0,0001
Medição da atividade CYP3A4
Os efeitos inibitórios de 40 e 80 nm nu e PC AuNP na atividade de CYP3A4 foram caracterizados. Como mostrado na Fig. 5, o BPEI-, LA- e PEG-AuNP de 40 nm e o BPEI-AuNP de 80 nm em LC 50 os valores inibiram a atividade catalítica do CYP3A4 em células C3A com a atividade correspondente de 20,1 a 31,4% em relação aos controles, independentemente do tamanho e da carga superficial. Concentrações atóxicas de LA- 80 nm e PEG-AuNP também suprimiram sua atividade (31,4 e 26,6%, respectivamente). No entanto, o PC melhorou amplamente a inibição de CYP3A4 induzida por AuNP de 40 e 80 nm, além de PEG-AuNP de 40 e 80 nm, mostrando atividade de 63 e 46% em comparação com os controles. Isso é consistente com estudos in vitro com tecido hepático humano e hepatócitos que o ácido tânico aniônico-AuNP e o BPEI-AuNP catiônico de 40 e 80 nm inibiram substancialmente a atividade catalítica do CYP3A4 [7, 25]. Em contraste, a expressão de mRNA induzida por PEI-AuNP catiônica e polivinilpirrolidona-AuNP neutra de CYP1A2, CYP2C9 e CYP3A4 em células HepG2 e CYP2B e CYP3A em fatia de fígado de rato, respectivamente [36, 37]. Um estudo recente relata que o BPEI-AuNP de 40 e 80 nm nu e PC BPEI-AuNP suprimiu substancialmente a atividade de CYP3A4 em hepatócitos humanos por meio de uma mudança conformacional na proteína ou bloqueando a bolsa de substrato como uma inibição reversível [7].
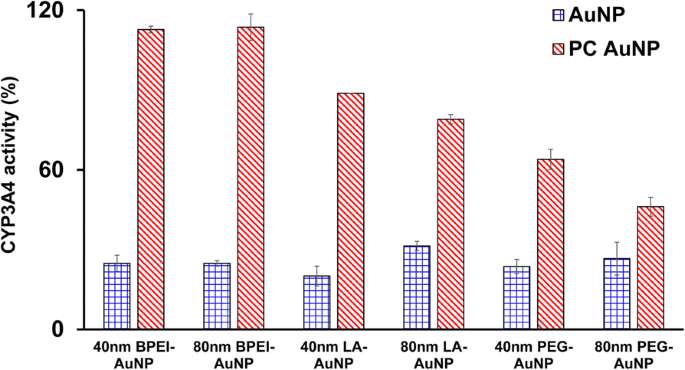
Efeito inibitório do AuNP na atividade do CYP3A4 em células C3A expostas ao BPEI-, LA- e PEG-AuNP de 40 e 80 nm na ausência e presença de PC por 24 h. Os valores representam a média ± D.P. ( n =3). BPEI polietilenimina ramificada, LA ácido lipóico, PEG polietilenoglicol, PC corona de proteína plasmática humana
Perfil de expressão gênica focada na via de toxicidade do PEG-AuNP de 40 nm
Dos genes representativos cobrindo 13 vias diferentes de estresse e toxicidade, um total de 212 genes (↓ 186 e ↑ 26 genes) foi diferencialmente expresso no LC 50 valor do PEG-AuNP de 40 nm (Fig. 6, Arquivo adicional 1:Tabelas S2 – S7). Os 12,3% (26 genes, ↓ 26, ↑ 0 genes) do total de genes (212 genes) estavam predominantemente envolvidos na β-oxidação do ácido graxo mitocondrial; para apoptose 11,3% (24 genes, ↓ 18, ↑ 6 genes); para danos ao DNA e via de reparo 11,3% (24 genes, ↓ 18, ↑ 6 genes); e para resposta ao choque térmico 11,3% (24 genes, ↓ 22, ↑ 2).
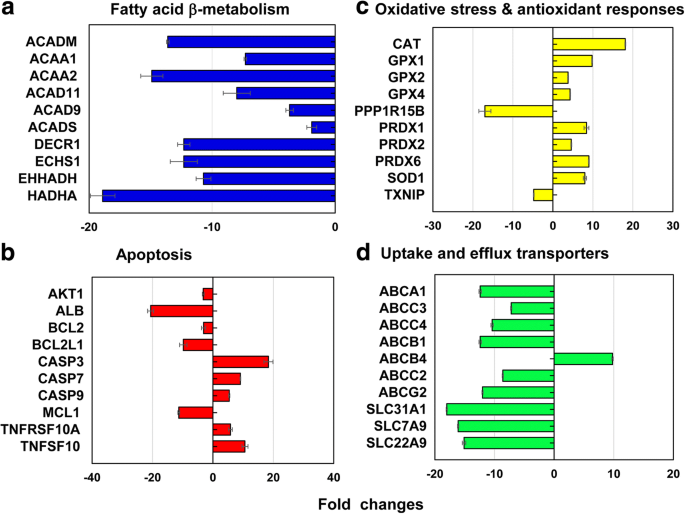
Genes representativos envolvidos em a oxidação de ácido graxo mitocondrial β, b apoptose, c estresse oxidativo e respostas antioxidantes, e d captação hepática e transportadores de efluxo em LC 50 valor de 40 nm PEG-AuNP. Todos os dados tiveram uma alteração de dobra <- 2 e> 2 em p <0,05. A análise da ontologia genética está listada no arquivo adicional 1:Tabelas S2 – S7
Na via de oxidação β do ácido graxo mitocondrial, genes que codificam três enzimas diferentes envolvidas na produção de acil-CoA e equivalentes redutores de NADH e FADH 2 foram suprimidos principalmente; Genes ACAD11, ACAD9, ACADM e ACADS em Acil-CoA desidrogenases (2,0 a 13,6 vezes); ACAA1 e ACAA2 em tiolases cetoacil-CoA (7,3 a 14,9 vezes); DECR1, ECHS1, EHHADH e HADHA (10,7 a 18,9 vezes) em enoil-CoA hidratase (Fig. 6a, Arquivo adicional 1:Tabela S2). A oxidação β do ácido graxo mitocondrial desempenha um papel importante na produção de acil-CoA e equivalentes redutores de NADH e FADH 2 , que está associada a quatro enzimas principais (acil-CoA desidrogenases, enoil-CoA hidratases, hidroxiacil-CoA desidrogenases e cetoacil-CoA tiolases [38, 39]. Além disso, transportadores de elétrons, NADH e FADH 2 , estão envolvidos no ciclo do ácido tricarboxílico (TCA) e na cadeia respiratória mitocondrial, resultando na produção de ATP. No estudo atual, o PEG de 40 nm induziu disfunção mitocondrial, uma perda de manutenção de ATP por meio de uma diminuição nos níveis intracelulares de ATP e FADH 2 , conseqüentemente definindo sua citotoxicidade em células C3 (Fig. 2a). O fenômeno semelhante foi relatado em hepatócitos humanos, HUVEC e HPTC, expostos ao BPEI-AuNP de 40 nm, indicando que a disfunção mitocondrial pode ser um mecanismo comum de toxicidade de AuNP, independentemente da carga superficial e tipos de células [7,8,9]. Um estudo recente relatou que a citotoxicidade relevante para a disfunção mitocondrial foi observada em células epiteliais de câncer de próstata imortalizadas e células epiteliais de câncer de pulmão em resposta a um inibidor da fosforilação de STAT3, OPB-51602 [40].
Na via de apoptose, os seis genes pró-apoptóticos de CASP3, CASP7, CASP9, TNFRSF10A, TNFRSF10B e TNFSF10 foram regulados positivamente, enquanto os seis genes anti-apoptóticos de AKT1, ALB, BCL2, BCL2L1, MCL1 e XIAP foram regulados para baixo (Fig. . 6b, Arquivo adicional 1:Tabela S3), que foi correlacionado com citotoxicidade dependente da dose em células C3A (Fig. 2a). No ponto de verificação de dano e reparo de DNA, os genes das quinases de checkpoint (CHEK1 / 2), os genes de reparo por excisão de DNA (ERCC1 / 2/3) e a ligase IV de DNA (LIG4) foram regulados positivamente, mas outros genes de reparo por excisão (ERCC5 / 6, XRCC1 / 5), o checkpoint quinase (CDKN1A) e os genes das proteínas quinases (PRKDC) foram regulados negativamente (2 a 19 vezes). Estes resultados sugeriram que a interferência induzida por PEG-AuNP de 40 nm com o ciclo celular e sistema de reparo de DNA pode se correlacionar com uma indução de morte celular em células C3A (Fig. 2a, Arquivo adicional 1:Tabela S3). Os genes que codificam duas proteínas de choque térmico diferentes (HSP) (A1A e A1B) foram regulados positivamente (10,2 a 14,2 vezes), mas as subfamílias A, B e C de HSP40; HSP90 membro 1; e HSP60 foram regulados para baixo (2 a 16 vezes) (Arquivo adicional 1:Tabela S4).
No estresse oxidativo e na resposta antioxidante, o PEG-AuNP de 40 nm em LC 50 genes de antioxidantes induzidos por valor e pró-oxidantes suprimidos, o que foi associado a uma diminuição na geração de ROS / RNS sendo o próprio antioxidante (Fig. 4b). Em genes antioxidantes, glutationa peroxidase (GPX) 1, GPX2, GPX4, PRDX1, PRDX2, PRDX6, superóxido dismutase (SOD) 1 e CAT foram induzidos (3,8 a 18,1 vezes). Em genes pró-oxidantes, TXNIP e PPP1R15B foram suprimidos (4,8 e 17 vezes, respectivamente) (Fig. 6c, Arquivo adicional 1:Tabela S5). Isso é consistente com um estudo anterior de que AuNP exibiu citotoxicidade induzida por estresse oxidativo em HepG2 e hepatócitos humanos, independentemente do tamanho [7, 19].
No metabolismo da fase I, os genes CYP3A4 e ESD foram amplamente suprimidos (7 vezes e 12 vezes, respectivamente). Especialmente, o efeito inibitório de PEG-AuNP de 40 nm na expressão de CYP3A4 foi correlacionado com uma diminuição na atividade de CYP3A4 (Fig. 5). Estudos recentes relataram que o BPEI-AuNP de 40 nm inibiu a expressão gênica de CYP1A2, CYP2C9 e CYP3A4 em hepatócitos humanos; ESD em HUVEC; e CYP1A1 em HPTC [7,8,9]. O estudo epidemiológico demonstrou que as enzimas CYP no tecido hepático de pacientes com CHC foram substancialmente inibidas pelo processo tumorigênico a nível molecular e funcional [41].
Perfil da expressão gênica do transportador de efluxo e captação de drogas
O desenvolvimento de multirresistência (MDR) por células tumorais é uma das principais causas de falhas no tratamento do câncer [42, 43]. A diminuição da captação de drogas mediada por transportadores de membrana integrais e o aumento do efluxo de drogas, incluindo P-glicoproteína (P-gp) e proteína de resistência ao câncer de mama (BCRP), é um dos principais mecanismos de MDR.
A expressão diferencial de genes de transportadores de efluxo e absorção de drogas em células C3A expostas a 40 nm PEG-AuNP mostrou que um total de 14 genes de transportadores ABC (↓ 12 e ↑ 2 genes) e um total de 21 genes de transportadores SLC (↓ 21 e ↑ 0 genes) foram substancialmente modulados em LC 50 valor (Figs. 6d e 7, Arquivo adicional 1:Tabela S7). Em transportadores de efluxo de drogas da família ABC, genes de proteínas associadas à resistência a múltiplas drogas (MRP3 / ABCC3), MRP4 (ABCC4) e proteína regulada de efluxo de colesterol (CERP / ABCA1) na membrana basolateral foram regulados para baixo (7,2 a 10,4 vezes). Os genes que codificam P-gp (ABCB1), MRP2 (ABCC2), BCRP (ABCG2) e esterolina 2 (ABCG8) em transportadores de efluxo canalicular também foram suprimidos (8,6 a 13,8 vezes). Em contraste, a multirresistência (MDR4 / ABCB4) na membrana canalicular e o transportador ABC de mitocôndria (MTABC3 / ABCB6) na membrana mitocondrial externa foram altamente reguladas positivamente (9,8 vezes e 5,8 vezes, respectivamente). Em transportadores de absorção de drogas, genes da proteína transportadora de cobre (CTR1 / SLC31A1) e em menor grau de transporte de ânions orgânicos (OAT7 / SLC22A9) também foram inibidos (18 vezes e 15 vezes, respectivamente). Estes resultados suportam um estudo recente que o BPEI-AuNP de 40 nm regula negativamente o MDR3 em hepatócitos humanos, mas regula positivamente o MRP3 em HUVEC, indicando a carga superficial e a interação dependente do tipo de célula entre AuNP e transportadores de efluxo [7, 8]. O estudo epidemiológico demonstrou que uma alta expressão de BCRP e uma baixa expressão de OCT3 ocorreu no tumor HCC, o que estava intimamente associado com a progressão do tumor e seu tamanho [44]. Um estudo anterior exibiu que o inibidor da P-gp, verapamil, aumentou a citotoxicidade da glutationa-AuNP conjugada com doxorrubicina em linhas de células de fibrossarcoma felino por meio do aumento da concentração intracelular da droga [45]. O estudo atual enfatiza que as informações derivadas dos mecanismos no PEG-AuNP de 40 nm identificaram uma ação separada, mas ainda complementar, na β-oxidação do ácido graxo mitocondrial, ciclo do TCA e cadeia respiratória, efluxo de drogas e transportadores de absorção, bem como a atividade do CYP3A4 em Células C3A (Fig. 7). Para o fim, isso irá destacar a interação do AuNP com os principais processos biológicos e seu mecanismo molecular subjacente no CHC, que pode estar ainda mais implicado no desenvolvimento de um alvo terapêutico mais eficaz no tratamento do CHC.
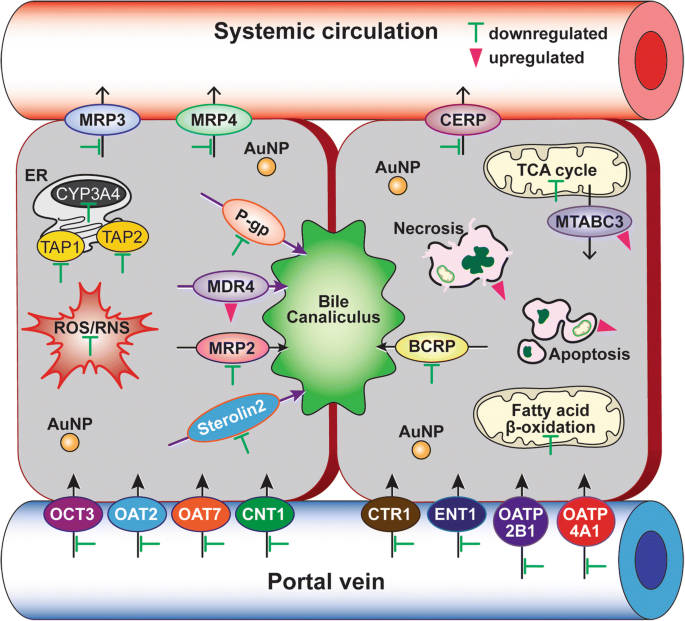
Uma representação esquemática dos mecanismos básicos de ação do PEG-AuNP 40 nm no tratamento do CHC. Barras verdes (uma inibição) e triângulos rosa (uma indução) indicam as vias e marcadores biológicos modificados com PEG-AuNP de 40 nm. A análise da ontologia genética está listada no arquivo adicional 1:Tabelas S2 – S7
Para validar a análise de expressão gênica de RT 2 array, os nove genes foram selecionados para PCR em tempo real. No arquivo adicional 1:Tabela S1, todos os nove genes foram modulados em LC 50 do PEG-AuNP de 40 nm. Essas mudanças transcricionais foram consistentes com as da análise de expressão gênica com matrizes de PCR (Fig. 6, Arquivo adicional 1:Tabelas S2-S7).
Conclusões
Apresentamos que a interação catiônica de BPEI-, LA- aniônica ou PEG-AuNP neutro com a proteína de plasma humano corona (PC) causou as mudanças em D H , PDI e potencial z de AuNP e influenciaram ainda mais as respostas celulares em células C3A. Todos os AuNP de 40 e 80 nm nus (sem PC) foram citotóxicos para células C3A além do LA-AuNP de 80 nm, mas o PC melhorou completamente suas citotoxicidades além do BPEI-AuNP de 80 nm. O BPEI-AuNP nu de 40 nm mostrou a maior captação celular seguido pelo PEG-AuNP de 40 nm e depois pelo LA-AuNP de 40 nm, enquanto o PC suprimiu a captação de AuNP além do PEG-AuNP de 80 nm. O BPEI-AuNP de 40 nm causou respostas bifásicas de estresse oxidativo (pró e antioxidante) em células C3A, enquanto o PEG-AuNP de 40 nm foi antioxidante. A atividade do CYP3A4 foi amplamente suprimida por todo o AuNP nu, independentemente do tamanho e das cargas de superfície, enquanto o PC melhorou substancialmente o seu efeito inibitório na atividade da enzima além do PEG-AuNP de 40 e 80 nm. Genes expressos diferencialmente em LC 50 valor de 40 nm PEG-AuNP estiveram principalmente envolvidos na β-oxidação de ácidos graxos mitocondriais e em um menor grau de transportadores de efluxo / captação hepática. O PEG-AuNP de 40 nm inibiu três enzimas principais na β-oxidação (acil-CoA desidrogenase, enoil-CoA hidratase e cetoacil-CoA tiolase), outras enzimas no ciclo do TCA e a cadeia respiratória mitocondrial para a produção de ATP. O PEG-AuNP de 40 nm aumentou a expressão de genes pró-apoptóticos e diminuiu genes anti-apoptóticos no LC 50 valor. Um alto nível de antioxidantes e um baixo nível de genes pró-oxidantes foram observados em células C3A expostas a 40 nm de PEG-AuNP. Além disso, genes de transportadores de efluxo e absorção de drogas localizados nas membranas basolateral e canalicular foram substancialmente modulados.
Abreviações
- ANOVA:
-
Análise de variância unilateral
- AuNP:
-
Nanopartículas de ouro nuas:sem PC
- BPEI:
-
Polietilenimina ramificada
- CYP:
-
Citocromo P450
- D H :
-
Diâmetros hidrodinâmicos
- DLS:
-
Espalhamento de luz dinâmico
- EDTA:
-
Ácido etilenodiaminotetracético
- EMEM:
-
Meio essencial mínimo de Eagle
- HCC:
-
Carcinoma hepatocelular humano
- HPTC:
-
Células tubulares renais humanas proximais
- HSD:
-
Teste de diferença significativa honesto de Tukey
- HUVEC:
-
Células da veia umbilical humana
- ICP-MS:
-
Espectrometria de massa com Plasma indutivamente acoplado
- LA:
-
Ácido lipóico aniônico
- LC 50 :
-
Concentração letal mediana
- MDR:
-
Resistência a múltiplas drogas
- NP:
-
Nanopartículas
- PBS:
-
Salina tamponada com fosfato
- PC:
-
Corona de proteína plasmática humana
- PDI:
-
Índice de polidispersidade
- PEG:
-
Polietilenoglicol neutro
- RNAi:
-
Interferência de RNA
- RNS:
-
Espécies reativas de nitrogênio
- ROS:
-
Espécies que reagem ao oxigênio
- SO:
-
Superóxido
- TEM:
-
Microscopia eletrônica de transmissão
- TFF:
-
Filtragem de fluxo tangencial
Síntese e estudo in vitro de uma sonda de modo duplo que direciona a integrina αvβ3
Deposição precisa in situ modificada em campo elétrico de fibras de cola médica eletroferadas no fígado para hemostasia rápida
Nanomateriais
- Manutenção de hotéis e seu impacto em seus resultados financeiros
- Nanobiossensor de ouro com base na ressonância de plasma de superfície localizada é capaz de diagnosticar a brucelose humana, apresentando um método rápido e acessível
- Síntese e atividade de oxidação de CO de 1D óxido binário misto CeO2-LaO x catalisadores de ouro suportados
- Síntese de aquecimento de estado sólido de composto de poli (3,4-etilenodioxitiofeno) / ouro / grafeno e sua aplicação para determinação amperométrica de nitrito e iodato
- Promoção do crescimento celular SH-SY5Y por nanopartículas de ouro modificadas com 6-mercaptopurina e um peptídeo penetrador de neurônio
- Anticorpo monoclonal de heparanase marcado com nanopartículas de ouro magnético e sua aplicação subsequente para imagens de ressonância magnética de tumor
- Efeitos do fulereno C60 na interação de difenil-N- (tricloroacetil) -amidofosfato com DNA em silico e sua atividade citotóxica contra linha celular leucêmica humana in vitro
- Avaliação de toxicidade de nanopartículas de PEG-PCCL e investigação preliminar sobre seu efeito antitumoral de carregamento de paclitaxel
- Influência dos nanotubos de carbono e seus derivados nas células tumorais in vitro e parâmetros bioquímicos, composição do sangue celular in vivo
- Avaliação da atividade do citocromo P450 3A4 inibido por nanopartículas de ouro e dos mecanismos moleculares subjacentes à sua toxicidade celular na linha celular C3A do carcinoma hepatocelular hu…