


Estabilidade e variabilidade de auto-montagem das proteínas da casca do microcompartimento bacteriano em resposta às mudanças ambientais
Resumo
Os microcompartimentos bacterianos (BMCs) são organelas protéicas que se auto-organizam e estão amplamente distribuídas no reino procariótico. Ao segmentar as principais enzimas metabólicas e vias usando uma camada poliédrica, os BMCs desempenham papéis essenciais na assimilação do carbono, na patogênese e na ecologia microbiana. O shell BMC é composto por vários homólogos de proteínas que se automontam para formar a arquitetura definida. Há um enorme interesse na engenharia de BMCs para desenvolver novos nanobiorreatores e estruturas moleculares. Aqui, relatamos a caracterização quantitativa da dinâmica de formação e automontagem de proteínas de casca BMC em pH variável e condições de sal usando microscopia de força atômica de alta velocidade (HS-AFM). Mostramos que a concentração de sal de 400 mM é propensa a resultar em manchas maiores de camada única formadas por hexâmeros de concha, e uma taxa dinâmica mais alta de automontagem do hexâmero foi observada em pH neutro. Também visualizamos a variabilidade de proteínas de shell de conjuntos hexaméricos a matrizes semelhantes a fibras. Este estudo avança nosso conhecimento sobre a estabilidade e variabilidade das automontagens da proteína BMC em resposta às mudanças microambientais, o que informará o projeto racional e a construção de estruturas de BMC sintéticas com a capacidade de remodelar sua automontagem e robustez estrutural. Ele também oferece uma caixa de ferramentas poderosa para avaliar quantitativamente a automontagem e a formação de nanoestruturas baseadas em BMC em aplicações de biotecnologia.
Introdução
Os microcompartimentos bacterianos (BMCs) são organelas proteicas, estruturalmente semelhantes às cápsides virais, que dividem o citoplasma das bactérias [1]. Eles são amplamente difundidos entre os filos bacterianos [2] e permitem que as bactérias compartimentem as principais vias metabólicas na ausência de organelas ligadas à membrana encontradas em eucariotos [3, 4]. BMCs são formados por um invólucro de proteína semipermeável que encapsula um núcleo de enzima luminal. A casca é composta por três tipos de componentes de proteínas estruturais, incluindo BMC-H (contendo um domínio Pfam00936), BMC-T (contendo dois domínios Pfam00936) e BMC-P (com um domínio Pfam03319) [5,6,7, 8,9]. Os principais componentes da casca são BMC-H, que aparecem como hexâmeros com superfícies convexas e côncavas e revestem as facetas da casca com seu lado côncavo voltado para fora [10] (Fig. 1). O BMC-P forma pentâmeros que são propostos para cobrir os vértices da forma icosaédrica, e o BMC-T forma pseudohexâmeros que estão localizados nas facetas da casca, presumivelmente responsáveis pela permeabilidade da casca.
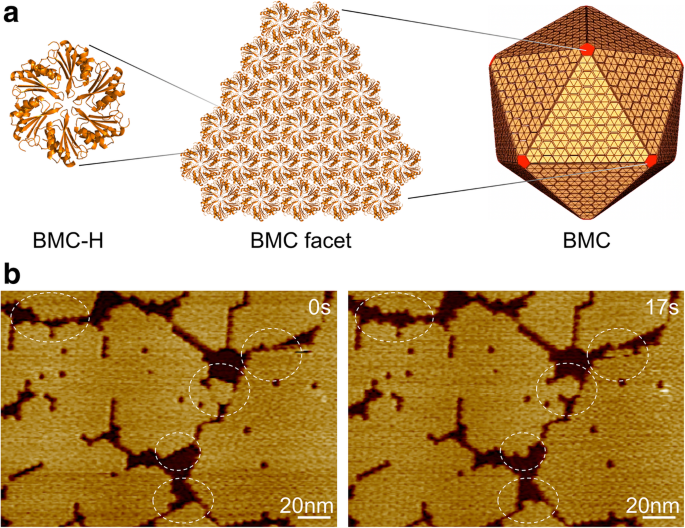
Microcompartimento bacteriano, organização da casca e automontagem. a Centenas de cópias de homólogos de proteínas do shell BMC se automontam para formar uma organela de proteína icosaédrica. As proteínas BMC-H, em amarelo, formam as facetas e as proteínas BMC-P, em vermelho, ocupam os vértices. b Topógrafos AFM de facetas de concha compostas por hexâmeros Hoch_5815 BMC-H. Eventos dinâmicos (círculos) foram observados em segundos usando HS-AFM
As interações proteína-proteína específicas garantem a automontagem das proteínas BMC para formar arquiteturas altamente definidas para cumprir sua funcionalidade metabólica. As interações laterais entre as proteínas da casca são consideradas o principal fator para determinar as propriedades de automontagem da casca icosaédrica [10]. Foi observado que homólogos de BMC-H podem formar várias formas, incluindo folhas bidimensionais [11, 12], nanotubos [13,14,15,16,17] e estruturas de filamentos [15, 18,19,20] .
Com base na automontagem, permeabilidade seletiva e propriedades de encapsulação enzimática das organelas que ocorrem naturalmente, os BMCs têm sido considerados como um sistema ideal com grande potencial em bioengenharia, incluindo a construção bioinspirada de biorreatores em nanoescala por encapsulamento de enzimas metabólicas e geração de novos andaimes moleculares com novas funções [21,22,23,24,25,26]. No entanto, algumas questões-chave ainda precisam ser abordadas na bioengenharia de BMC, por exemplo, quão estáveis as estruturas de BMC são e como manipular e avaliar efetivamente a automontagem e formação de agregados de proteína BMC. Investigações das estruturas e montagem de cascas de BMC e BMCs inteiras foram realizadas usando cristalografia de raios-X, microscopia eletrônica (EM), microscopia de fluorescência e espalhamento dinâmico de luz (DSL) [10, 11, 16, 22, 27,28 , 29,30,31]. Recentemente, exploramos o AFM de alta velocidade (HS-AFM) para conduzir a primeira visualização do processo de automontagem dinâmica das proteínas BMC-H [12].
Neste trabalho, usamos HS-AFM para monitorar a dinâmica estrutural de patches BMC-H sob variados pH e condições iônicas, o que fornece uma visão sobre a modulação da montagem da proteína shell BMC e oferece uma ferramenta poderosa para avaliação quantitativa, na resolução molecular , na estabilidade e variabilidade da auto-montagem da proteína de casca de BMC.
Métodos
Preparação da amostra
A proteína BMC-H purificada (Hoch_5815) de Haliangium ocraceum foi gentilmente cedido pelo Dr. Kerfeld (Laboratório Nacional Lawrence Berkeley). Para troca de tampão, amostras de estoque em ~ 80 mg mL −1 em tampão Tris (50 mM Tris-HCl, pH 7,8, 100 mM KCl, 10 mM MgCl 2 ) foram diluídos para 0,5 mg mL −1 usando o buffer desejado antes da imagem AFM (arquivo adicional 1:Figura S1). O tampão de controle é Tris-HCl 50 mM (pH 7,8) e MgCl 10 mM 2 .
Microscopia de força atômica
Os tampões desejados foram usados para a absorção da amostra em imagens de mica e AFM. Após 5 min de absorção na mica, Hoch_5815 foram enxaguados com o tampão desejado para remover as proteínas imobilizadas e, em seguida, imagens usando AFM (arquivo adicional 1:Figura S1). Imagens HS-AFM foram capturadas a 30 ou 40 Hz em solução em modo AC usando um JPK NanoWizard ULTRA speed AFM equipado com um scanner ULTRA Speed 2.8 μm e sondas Ultra-Short Cantilever USC-0.3 MHz (NanoWorld). Forças de carregamento mínimas de ~ 100 picoNewton foram aplicadas durante a imagem AFM para reduzir a perturbação da montagem da proteína [12, 32,33,34,35,36].
Processamento e análise de imagem
A análise de imagens foi inicialmente realizada usando JPK SPM Data Processing (JPK). A análise de imagem HS-AFM foi realizada usando uma macro customizada no Image SXM (http://www.ImageSXM.org.uk), conforme descrito anteriormente [12]. Para analisar os tamanhos dos patches Hoch_5815, imagens de 512 × 512 pixels capturadas a uma taxa de varredura de 30 Hz foram achatadas para remover qualquer inclinação XY e limite Z, seguido por conversão binária para exibir proteína versus não proteína. A análise de partículas foi usada para calcular a área de superfície das proteínas nessas imagens binárias. Patches foram definidos como objetos separados por> 3 pixels (~ 2 nm), a fim de identificar patches individuais versus patches adjacentes. Os testes iniciais mostraram que, se um número maior de pixels for definido, os patches adjacentes podem ser contados como um único patch contínuo, ao passo que, usando um número de pixel menor, as lacunas entre hexâmeros individuais nos patches podem ser contadas incorretamente como o limite entre os patches. Para analisar a dinâmica da proteína, séries de imagens de 256 × 256 pixels capturadas na velocidade de varredura de 40 Hz foram analisadas dando uma resolução temporal de aproximadamente 6,4 s por quadro. Imagens binárias foram subtraídas da imagem anterior na série para mostrar imagens AFM diferentes. A análise de partículas das imagens de diferença foi empregada para contar a área de proteínas montadas e desmontadas. A equação usada para calcular a taxa dinâmica é mostrada a seguir:
$$ \ mathrm {Taxa} \ \ mathrm {de} \ \ mathrm {dinâmico} \ \ mathrm {eventos} \ (R) =\ frac {\ mathrm {Número} \ \ mathrm {de} \ \ mathrm {hexâmeros} \ \ mathrm {adicionado} \ \ mathrm {ou} \ \ mathrm {removido} \ \ mathrm {in} \ \ mathrm {a} \ \ mathrm {s} \ mathrm {eries} \ \ mathrm {of} \ \ mathrm {frame} \ mathrm {s} \ kern0.5em (N)} {\ mathrm {Total} \ \ mathrm {surface} \ \ mathrm {area} \ \ mathrm {of} \ \ mathrm {protein} \ \ mathrm { in} \ \ mathrm {frame} \ kern0.5em (A) \ times \ mathrm {time} \ kern0.5em (T)}, $$
onde N representa a soma de pixels brancos e pretos em uma imagem de diferença limiar dividida pelo número de pixels correspondentes a um único hexâmero naquela escala (Arquivo adicional 1:Figura S3, Figura S5). Os dados são apresentados como média ± desvio padrão (DP). A análise estatística foi realizada usando ANOVA multivariada ou ANOVA de dois fatores, conforme especificado.
Resultados
Usamos as proteínas BMC-H (Hoch_5815) de um myxobacterium Haliangium ocraceum , que foram expressos em Escherichia coli e caracterizados como hexâmeros com simetria sêxtupla [12]. Os hexâmeros Hoch_5815 podem se automontar para formar folhas de camada única na segunda escala de tempo, que representam os componentes estruturais básicos da arquitetura icosaédrica BMC (Fig. 1a). A imagem HS-AFM nos permite visualizar a montagem dinâmica e a flexibilidade organizacional dos fragmentos de folha (Fig. 1b) e estimar quantitativamente o tamanho do patch e a taxa dinâmica das proteínas BMC-H usando a análise de imagem desenvolvida (consulte a seção "Métodos").
Resposta à variação de pH
Medimos as mudanças no tamanho do patch como uma indicação da capacidade geral do Hoch_5815 de se automontar. O tamanho do remendo aumenta com o aumento do pH de 3 para 10 (Fig. 2a; Arquivo adicional 1:Figura S2, Tabela S1), sugerindo que pH alto é mais favorável para a automontagem de proteínas Hoch_5815 do que condições de pH baixo. Isso é um tanto distinto dos comportamentos de montagem das proteínas RmmH, que foram consideradas insolúveis em pH 6, formam matrizes ordenadas de nanotubos em pH 8 e eram propensas a se desmontar em pH 10 [13]. Além disso, observamos um alto grau de variabilidade estrutural das automontagens HOCH_5815 (conforme indicado por um grande SD na Fig. 2a, Arquivo adicional 1:Figura S2).
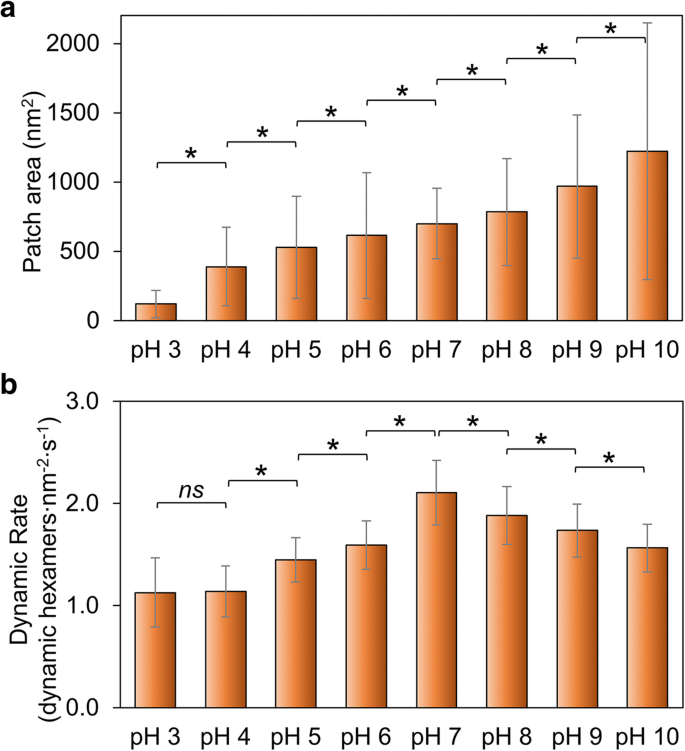
Efeitos do pH ambiental na automontagem de Hoch_5815. a As áreas de superfície médias de manchas individuais de Hoch_5815 determinadas por AFM ( n =50) (Arquivo adicional 1:Figura S2). b As taxas médias de eventos dinâmicos determinadas por HS-AFM ( n =50). * p <0,05, ns não significativo (ANOVA multivariada)
A imagem AFM na automontagem de proteínas Hoch_5815 em folhas de revestimento revelou que a formação de folhas de revestimento é atribuída a uma combinação da montagem e desmontagem de hexâmeros [12]. Examinamos ainda as taxas de dinâmica de auto-montagem Hoch_5815 e eventos dinâmicos sob diferentes pH (arquivo adicional 1:Tabela S2) para explorar a estabilidade das interações proteína-proteína Hoch_5815. A taxa de dinâmica de automontagem é mais alta em pH 7 e diminui em condições ácidas e alcalinas (Fig. 2b; Arquivo adicional 1:Figura S3). Em particular, diminui rapidamente em condições ácidas, notavelmente de pH 7 para pH 6 e parece relativamente constante entre pH 4 e pH 3, como mostrado na Fig. 2b.
É provável que o pH tenha um grande impacto nas propriedades eletrostáticas dos resíduos de aminoácidos localizados na interface hexâmero-hexâmero. A dinâmica diminuída e um tamanho menor de remendos de casca observados em condições ácidas ilustram que Hoch_5815 tem uma capacidade de automontagem reduzida. A dinâmica reduzida e um tamanho maior de manchas de casca observados nas condições alcalinas sugerem interações hexâmero-hexâmero estáveis, enquanto a dinâmica aumentada de hexâmeros Hoch_5815 implica interações hexâmero-hexâmero flexíveis na condição de pH neutro.
Resposta à variação das concentrações de sal
Também verificamos se a concentração de sal do buffer tem impactos na montagem de Hoch_5815. Em baixas concentrações (100–200 mM) de MgCl 2 , CaCl 2 e KCl, as proteínas Hoch_5815 formam manchas relativamente menores do que aquelas montadas em concentrações mais altas (300-500 mM) (Fig. 3a; Arquivo adicional 1:Figura S4). A 500 mM, observamos folhas Hoch_5815 de camada dupla ou múltipla (arquivo adicional 1:Figura S4). Essas observações são consistentes com a descoberta anterior de que maior força iônica poderia facilitar a formação de cristais 2D mais extensos e bem ordenados por CcmK, as proteínas da casca dos carboxissomos para assimilação de carbono [37]. No entanto, os nanotubos altamente ordenados formados por RmmH foram desmontados quando a concentração de NaCl foi aumentada de 50 para 500 mM [13], indicando os mecanismos potencialmente diferentes que medeiam a formação de folhas planas e formatos tubulares por hexâmeros de casca.
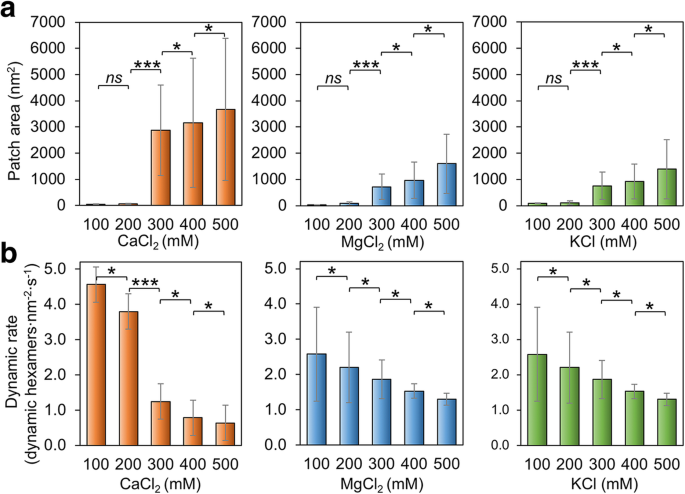
Efeitos da concentração de sal na automontagem de Hoch_5815. a As áreas médias do patch medidas por AFM em um intervalo de 100–500 mM CaCl 2 , MgCl 2 , e KCl ( n =50). O aumento na concentração de sal resultou em tamanhos de remendo aumentados. Mudanças significativas na área do remendo foram observadas entre 200 e 300 mM (*** p <0,001, * p <0,05, ns não significativo, ANOVA de dois fatores). b As taxas médias de eventos dinâmicos determinadas a partir de séries de imagens AFM de alta velocidade em um intervalo de 100–500 mM CaCl 2 , MgCl 2 , e KCl ( n =50). Cada mudança de 100 mM na concentração de sal levou a uma mudança significativa na taxa de eventos dinâmicos (*** p <0,001, * p <0,05, ns não significativo, ANOVA de duas vias)
Além disso, as variações da auto-montagem Hoch_5815 causadas pelas mudanças no MgCl 2 e as concentrações de KCl são relativamente semelhantes. Em contraste, a mudança no tamanho do patch é mais pronunciada (até um aumento de 3.000 vezes) quando o CaCl 2 a concentração é aumentada de 200 para 300 mM (Fig. 3a), sugerindo a maior sensibilidade da auto-montagem Hoch_5815 para CaCl 2 do que para MgCl 2 ou KCl.
A taxa dinâmica de auto-montagem Hoch_5815 também é afetada por mudanças na concentração de sal tampão. O aumento em MgCl 2 , CaCl 2 , ou as concentrações de KCl podem resultar no declínio da taxa dinâmica Hoch_5815 (Fig. 3b; Arquivo adicional 1:Figura S5). Dado o aumento no tamanho do patch observado sob altas concentrações de sal (Fig. 3a), parece que as interações laterais entre os hexâmeros Hoch_5815 são mais estáveis sob altas concentrações de sal. Mudanças no CaCl 2 a concentração teve uma resposta mais pronunciada e houve uma mudança significativa na taxa de eventos dinâmicos entre 200 e 300 mM (Fig. 3b), enquanto as respostas às mudanças no MgCl 2 e KCl são relativamente semelhantes, consistentes com as mudanças no tamanho do remendo (Fig. 3a). Curiosamente, as maiores proporções de eventos de montagem versus eventos de desmontagem foram observadas sob 400 mM de MgCl 2 , CaCl 2 ou KCl (Arquivo adicional 1:Tabela S2). Isso levou à formação de grandes e estáveis conjuntos Hoch_5815 de camada única sob 400 mM de sal (Arquivo adicional 1:Figura S4). Os conjuntos de camada dupla observados a 500 mM também são estáveis e exibem baixas taxas de movimento do hexâmero.
Flexibilidade do conjunto de proteína BMC-H
Ao reduzir a força de varredura para 100 pN, minimizamos os efeitos da varredura da ponta de AFM na montagem de proteínas BMC e obtivemos imagens de AFM de resolução molecular de hexâmeros individuais (Fig. 4). Ambos os eventos de montagem e desmontagem podem ser vistos na mesma visualização, verificando a natureza dinâmica das montagens do shell BMC ao invés dos artefatos de varredura da ponta [12]. A imagem HS-AFM também revelou a variabilidade das agregações da proteína Hoch_5815. Ao obter imagens das amostras em pH 7,5 na presença de apenas 10 mM MgCl 2 , surpreendentemente, ocasionalmente observamos a formação de estruturas semelhantes a fibras junto com a desmontagem dos hexâmeros Hoch_5815 na segunda escala de tempo (Fig. 4a). Essas estruturas semelhantes a fibras podem ser densamente empacotadas em paralelo, semelhantes aos feixes de nanotubos montados por hexâmeros de casca [13,14,15,16]. No entanto, o espaço entre duas fibras é 3,72 ± 0,31 nm ( n =30) e sua altura média é 2,46 ± 0,22 nm ( n =30), menos do que as folhas de casca formadas por hexâmeros Hoch_5815 (3,45 ± 0,16 nm, n =25) (Fig. 4b – f). Essas estruturas de fibra são bastante flexíveis e dinâmicas durante a geração de imagens e podem exibir arquiteturas retas ou em espiral com tamanhos diferentes. Dada a aparência concomitante das estruturas de fibra com a desmontagem de hexâmeros Hoch_5815 e sua altura reduzida em comparação com as folhas de hexâmero de camada única, especulamos que essas estruturas semelhantes a fibras são formadas pelos peptídeos Hoch_5815 individuais desmontados dos hexâmeros (Fig. 4g) . É provável que a absorção do substrato em condições tampão específicas (como baixa força iônica) possa levar à fixação dos lados alfa-hélice dos peptídeos Hoch_5815 à superfície do substrato e à ligação linear dos peptídeos com os peptídeos vizinhos, embora seja assumido que as interações intra-hexâmero são provavelmente fortes [5]. O mecanismo detalhado subjacente à variabilidade da agregação da proteína shell ainda precisa ser elucidado.
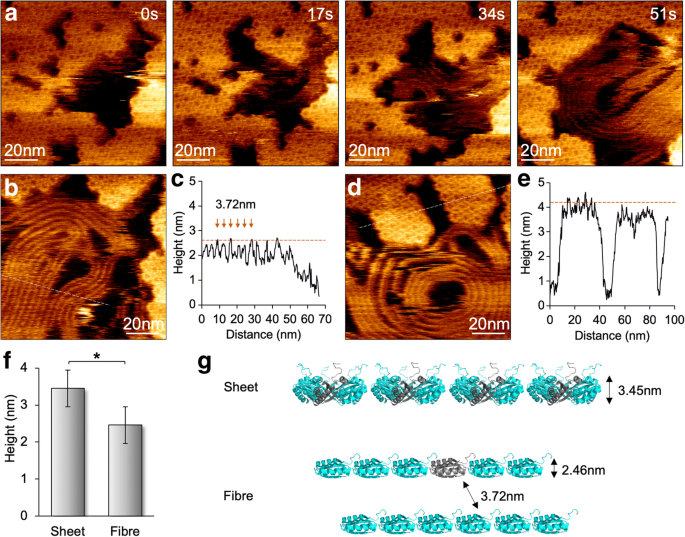
Formação e dinâmica de estruturas fibrosas junto com os conjuntos de folha de casca sob HS-AFM. a Aparência de estruturas semelhantes a fibras durante a desmontagem de folhas de concha compostas por hexâmeros Hoch_5815, conforme mostrado pela série de imagens AFM. b Topografia AFM de estruturas de fibra. c Análise de seção transversal (linha tracejada no painel b ) revela um espaçamento de 3,72 ± 0,31 nm ( n =30) entre estruturas de fibra adjacentes, e a altura média é 2,46 ± 0,22 nm ( n =30). d Topografia AFM de manchas de concha compostas por hexâmeros Hoch_5815. e Análise de seção transversal (linha tracejada no painel d ) revela que a altura média dos hexâmeros Hoch_5815 é 3,45 ± 0,16 nm ( n =25). f As estruturas semelhantes a fibra apresentam uma altura reduzida em comparação com folhas planas consistindo de hexâmeros Hoch_5815 (* p <0,05, ANOVA de dois fatores). g Organização proposta e formação da estrutura semelhante a fibra, representando uma cadeia de monômeros Hoch_5815
Discussão
BMCs compreendem centenas de proteínas que se auto-montam para formar as estruturas de ordem superior. O invólucro BMC, que consiste em vários homólogos de proteínas, é um sistema ideal para estudar a automontagem e as interações de proteínas. Como uma técnica poderosa para analisar a organização da biomembrana, montagem de proteínas e interações físicas que são altamente relevantes para os papéis fisiológicos dos sistemas biológicos [32, 35, 38, 39], AFM foi explorado para visualizar a organização e dinâmica de automontagem de Proteínas de shell BMC e as arquiteturas e características mecânicas das estruturas BMC [12, 30, 31, 40,41,42]. Este trabalho representa, até onde sabemos, a primeira determinação quantitativa da dinâmica de automontagem das proteínas da casca BMC na formação de folhas bidimensionais em resposta a mudanças ambientais usando AFM. Os resultados destacam a variabilidade inerente e a dependência ambiental da auto-montagem da proteína BMC-H. Comparado com EM e DSL, AFM exibe grande potencial no monitoramento das ações dinâmicas da auto-montagem da proteína BMC em tempo real com detalhes moleculares.
As interações proteína-proteína são de importância significativa na formação e modelagem do invólucro BMC [10]. A concentração de proteína também foi documentada como um fator crítico para conduzir a formação de casca [41, 43]. Além disso, estudos de solubilidade in vitro ilustraram que o pH e a força iônica em solução podem influenciar a estabilidade estrutural dos BMCs [17, 27], bem como os comportamentos de montagem das proteínas da casca do BMC na formação de folhas bidimensionais [37, 41] ], nanotubos [13, 17] e nanocages [28], uma reminiscência de seu impacto na montagem do capsídeo do vírus [44, 45]. Também encontramos precipitação de proteína e nenhuma mancha formada quando o pH> 10 e <3 ou a concentração de sal <10 mM ou> 600 mM (dados não publicados). Aqui, mostramos ainda que a tendência e a dinâmica de montagem dependem do pH e da concentração de sal. Embora as proteínas de casca possam se automontar em uma ampla faixa de pH, o ambiente de pH neutro parece ser capaz de aumentar a dinâmica de montagem (Fig. 2b). Verificou-se que cátions com concentração ≥ 300 mM promovem a formação de folhas bidimensionais; Catiões 400 mM parecem ser desejáveis para a formação de folhas grandes e estáveis de camada única (Fig. 3). Essas condições se alinham com as condições citosólicas das células bacterianas e são fisiologicamente relevantes. Por exemplo, na maioria das condições fisiologicamente relevantes, o pH de E. coli o citosol é de aproximadamente 7,4-7,8 [46] e a concentração de íons é de aproximadamente 100-400 mM, o que é vital para interações de proteínas, ligação proteína-ligante, sinalização, manutenção de potenciais eletrostáticos de membrana e gradiente de proteína através das membranas [47, 48]. Embora a forma como as interações entre as amostras e o substrato de mica afetam a automontagem das proteínas BMC ainda precisam ser investigados, a imagem AFM fornece a oportunidade para analisarmos quantitativamente as mudanças dinâmicas da automontagem da proteína BMC em resposta às variações ambientais.
A dinâmica de montagem dependente do ambiente de proteínas BMC na formação de fragmentos de casca descritos aqui pode representar seus comportamentos na formação de todo o BMC. Na verdade, as estruturas 3D BMC parecem ser organelas mantidas dinamicamente projetadas na natureza. BMCs apresentam notável flexibilidade estrutural e heterogeneidade; a suavidade mecânica das estruturas de shell BMC determinada por AFM nanoindentação [30] e a dinâmica de não-equilíbrio da montagem BMC revelada por simulações computacionais [49] destacou as diferenças entre BMC e conjuntos de vírus robustos. Da mesma forma, a biossíntese de carboxissomos foi elucidada para se correlacionar com luz e chaperons [50, 51]. Muito recentemente, foi indicado que CcmK3 e CcmK4 podem formar heterohexâmeros e cobrir a concha do carboxissomo de uma maneira dependente do pH, possivelmente fornecendo um meio para regular a permeabilidade da concha do carboxissomo e CO 2 assimilação no microambiente altamente dinâmico [52]. O mecanismo exato subjacente a como as condições ambientais em solução afetam a montagem termodinâmica das proteínas BMC ainda precisa ser investigado, por exemplo, usando uma combinação de estudos experimentais e simulações computacionais.
Dada a automontagem de estruturas de BMC, há um interesse significativo na engenharia de BMCs e no projeto de novos nanobiorreatores baseados em BMC, andaimes moleculares e biomateriais em aplicações de biotecnologia, por exemplo, melhorando o metabolismo celular, encapsulamento enzimático, entrega molecular e terapia . Conhecimento avançado sobre a resiliência estrutural e variabilidade de BMCs em resposta a mudanças ambientais não apenas informará estratégias para a produção de nanoestruturas baseadas em BMC robustas em hospedeiros heterólogos, ou seja, E. coli ou plantas [31, 53, 54], mas também pavimentam o caminho para modular a formação de nanomateriais 2D, bem como a abertura e o fechamento de gaiolas de proteína baseadas em casca de BMC, facilitando assim a regulação funcional e distribuição molecular direcionada. Anteriormente, demonstramos a viabilidade do uso de abordagem de modificação genética para manipular os contatos específicos nas interfaces das proteínas da casca e seus comportamentos de automontagem [12]. Este estudo fortalece nossa caixa de ferramentas para avaliar e manipular a automontagem do shell BMC em ambientes variados.
Conclusões
Em resumo, exploramos o HS-AFM para realizar as investigações quantitativas da auto-montagem da proteína shell BMC sob diferentes condições de pH e sal. A formação de remendos de camada única maiores de hexâmeros de concha mostrou ser promovida na concentração de sal de 400 mM, e o pH neutro resultou em uma taxa dinâmica mais alta de automontagem do hexâmero. A transição organizacional de proteínas de shell de conjuntos hexaméricos para matrizes semelhantes a fibras também foi visualizada. Este estudo ilustrou que as condições ambientais desempenham um papel importante na determinação da organização e automontagem das proteínas da casca BMC.
Abreviações
- BMC:
-
Microcompartimento bacteriano
- BMC-H:
-
Hexâmero de microcompartimento bacteriano
- BMC-P:
-
Pentâmero de microcompartimento bacteriano
- BMC-T:
-
Trímero de microcompartimento bacteriano
- DSL:
-
Espalhamento de luz dinâmico
- E. coli :
-
Escherichia coli
- EM:
-
Microscópio eletrônico
- HS-AFM:
-
Microscopia de força atômica de alta velocidade
Deposição de camada atômica aprimorada com plasma de baixa temperatura de SiO2 usando dióxido de carbono
Aprimoramento dielétrico de capacitores de Al2O3 / ZrO2 / Al2O3 MIM depositados na camada atômica por recozimento por micro-ondas
Nanomateriais
- Avaliando a IoT e o impacto do 5G
- Inovação em redes privadas:Configure e altere a largura de banda e a duração em tempo real
- Revelando a estrutura atômica e eletrônica das nanofibras de carbono de copo empilhado
- Estudo de primeiros princípios sobre a estabilidade e imagem STM de Borophene
- Síntese fácil de óxido de estanho mesoporoso semelhante a buraco de minhoca via auto-montagem induzida por evaporação e propriedades aprimoradas de detecção de gás
- Sondando as propriedades estruturais, eletrônicas e magnéticas dos aglomerados Ag n V (n =1–12)
- Robôs, cobots e automação:a pandemia está impulsionando uma onda de mudanças na fabricação?
- Como a recuperação da manufatura mudará o recrutamento e a retenção de funcionários
- O impacto ambiental da fabricação da revolução industrial à automação e tudo mais
- A automação e os robôs mudarão o mundo?