


F-actina regula a diferenciação osteoblástica de células-tronco mesenquimais em nanotubos de TiO2 por meio de MKL1 e YAP / TAZ
Resumo
Titânio e ligas de titânio são amplamente utilizadas em implantes ortopédicos. Modificar a nanotopografia fornece uma nova estratégia para melhorar a osseointegração de substratos de titânio. A polimerização da actina filamentosa (F-actina), como uma estrutura de carga mecânica, é geralmente considerada como estando envolvida na migração celular, endocitose, divisão celular e manutenção da forma celular. Se a F-actina está envolvida e como ela funciona na diferenciação osteogênica induzida por nanotubos de células-tronco mesenquimais (MSCs) ainda precisam ser elucidados. Neste estudo, nós fabricamos TiO 2 nanotubos na superfície de um substrato de titânio por oxidação anódica e caracterizou suas características por microscopia eletrônica de varredura (SEM), análise dispersiva de energia de raios-X (EDS) e microscopia de força atômica (AFM). A coloração com fosfatase alcalina (ALP), Western blotting, qRT-PCR e coloração de imunofluorescência foram realizadas para explorar o potencial osteogênico, o nível de F-actina e a expressão de MKL1 e YAP / TAZ. Nossos resultados mostraram que o diâmetro interno e a rugosidade do TiO 2 os nanotubos aumentaram com o aumento da voltagem de oxidação anódica de 30 para 70 V, enquanto sua altura era de 2 μm de forma consistente. Além disso, quanto maior o diâmetro do tubo, maior será a capacidade do TiO 2 nanotubos para promover a diferenciação osteogênica de MSCs. A inibição da polimerização de F-actina por Cyto D inibiu a diferenciação osteogênica de MSCs, bem como a expressão de proteínas contidas em complexos de adesão focal, como vinculina (VCL) e quinase de adesão focal (FAK). Em contraste, após o tratamento com Jasp, a polimerização da F-actina aumentou a expressão de RhoA e dos fatores de transcrição YAP / TAZ. Com base nesses dados, concluímos que TiO 2 nanotubos facilitaram a diferenciação osteogênica de MSCs, e essa capacidade foi aprimorada com o aumento do diâmetro dos nanotubos dentro de uma certa faixa (30-70 V). A F-actina mediou esse processo por meio de MKL1 e YAP / TAZ.
Introdução
O titânio e as ligas de titânio, devido à sua excelente biocompatibilidade, resistência à corrosão e propriedades mecânicas, são amplamente utilizadas em aplicações clínicas como substituições totais de articulações e implantes dentários [1,2,3]. No entanto, ainda existem muitos desafios a serem resolvidos, incluindo soltura asséptica e infecção [4, 5]. Nos últimos anos, foram realizados vários estudos com o objetivo de melhorar a osseointegração e as propriedades antibacterianas. Por exemplo, o MoS 2 / PDA-RGD revestimento em implantes de titânio pode não só promover a integração de um implante de titânio com o osso hospedeiro, mas também inibir o crescimento bacteriano com uma alta eficiência [6]. Além disso, a topografia da superfície tem atraído cada vez mais atenção, e a modificação topográfica difere das modificações químicas mudando apenas a estrutura em escala micro e nano. A estimulação de sinais químicos nas células é instável e citotóxica. Em comparação, sinais físicos seguros e controláveis podem evitar alguns efeitos colaterais causados por moléculas químicas. Portanto, a modificação topográfica da superfície dos implantes e a regulação da osseointegração através da estrutura topográfica podem fornecer uma nova forma de resolver o problema clínico da má osseointegração após o implante de próteses.
No campo da engenharia de tecido ósseo e regeneração óssea, a interação célula-morfologia é considerada uma estratégia de gerenciamento promissora para o controle preciso da função e diferenciação das células-semente. Ao mesmo tempo, o próprio osso tem uma hierarquia elegante dentro da faixa nanométrica e mícron [7]. Portanto, a morfologia da superfície pode fornecer um nicho semelhante, que pode imitar a estrutura óssea natural e promover a diferenciação osteogênica de células-tronco mesenquimais na superfície do osso hospedeiro e do implante. Morfologias de superfície podem consistir em muitas estruturas diferentes, incluindo nanotubos, nanofios, nanoporos e assim por diante. Em particular, os arranjos de nanotubos têm atraído amplo interesse em muitos campos nos últimos anos devido às suas características únicas de superfície, como alta proporção superfície-volume, plasticidade biológica e alta capacidade de adsorção. Por exemplo, um novo estudo mostra que os nanotubos de nitreto de boro (BNNT) constituem um material sensível a gás que pode ser usado como um sensor de gás para monitorar a operação de transformadores, detectando a composição e o conteúdo dos gases dissolvidos no óleo [8]. Na biomedicina, a topografia de superfície também é capaz de direcionar comportamentos celulares, incluindo migração, adesão, proliferação e diferenciação celular. Os estudos mais recentes revelam que a topografia em nanoescala pode direcionar as células-tronco mesenquimais (MSCs) para se diferenciarem em osteoblastos, de modo a reforçar a osseointegração precoce [9,10,11,12]. É ainda relatado que a modificação de superfície em micro e nanoescala combinada pode fazer com que as CTMs se diferenciem em células musculares lisas contráteis [13]. No entanto, os mecanismos moleculares de como a topografia de superfície direciona o destino das células ainda precisam ser elucidados, o que é importante para a avaliação da segurança do material e o design do material.
A (F) -actina filamentosa, também chamada de microfilamento, é um dos três principais componentes do citoesqueleto nas células eucarióticas. É composto por polímeros de globular (G) -actina, modificados por inúmeras outras proteínas. A F-actina tem polaridade estrutural devido ao fato de que todas as subunidades do microfilamento apontam para a mesma extremidade. A extremidade farpada é direcionada a um monômero adjacente diferente, enquanto a extremidade pontiaguda possui uma subunidade de actina com o local de ligação do ATP exposto. Ou seja, o ATP está envolvido no processo de transformação entre G-actina e F-actina. Este processo encontra-se em equilíbrio dinâmico, com polimerização e despolimerização ocorrendo simultaneamente, também conhecido como treadmilling, frequentemente visto em lamelipódios e filopódios [14]. Portanto, é óbvio que a dinâmica da actina desempenha um papel importante em funções celulares como migração celular, divisão celular e manutenção da forma celular. No entanto, a F-actina não apenas atua como uma estrutura física de suporte à carga mecânica, mas também participa de outros comportamentos biológicos, como transdução de sinal e expressão gênica. Evidências acumuladas demonstram que a F-actina pode converter sinais físicos em sinais químicos ao interagir com outras proteínas [15,16,17,18]. Por exemplo, a reconstrução biomecânica e geométrica promove a apoptose das células tumorais, evitando a sobreposição da polimerização do monômero de actina a F-actina [15]. Os pulsos de ultrassom aumentam a osteogênese das células-tronco mesenquimais humanas ao inibir a despolimerização da F-actina [16]. Nosso estudo anterior também mostrou que a deformação mecânica aumenta a estabilidade da F-actina [17]. Como a estrutura oca dos nanotubos fornece menos locais de adesão para as células, o rearranjo do citoesqueleto é inevitável para manter o equilíbrio biomecânico. Consequentemente, definitivamente temos razões para acreditar que a F-actina provavelmente medeia a diferenciação celular induzida por nanotopografia.
Neste estudo, nós fabricamos TiO 2 nanotubos, modificou sua topografia por oxidação anódica e explorou sua capacidade de promover a diferenciação osteogênica de MSCs. Em seguida, investigamos se a F-actina desempenha um papel crítico na mecanotransdução. A citocalasina D (Cyto D), que se liga competitivamente à extremidade farpada da F-actina para evitar que a G-actina se incorpore ao filamento, foi usada para inibir a polimerização da F-actina e o jasplacinolido (Jasp) foi usado para aumentar a estabilização do montagem de actina. Além disso, também queríamos elucidar como a F-actina funciona na mudança de pistas físicas em sinais bioquímicos. Com base nos resultados de nosso estudo anterior, formulamos a hipótese de que a via MAPK pode estar envolvida neste processo [17]. Fatores de transcrição, como proteína associada a Sim (YAP) / coativador transcricional com motivo de ligação de PDZ (TAZ) e MKL1, que são considerados mecanossensores e mecanotransdutores, também foram os sujeitos de nosso estudo para rastrear como a F-actina influencia as células-tronco destino, porque alguns estudos em outras áreas implicaram que eles estavam relacionados à F-actina [19,20,21]. De forma geral, esperamos esclarecer o papel da F-actina no processo de diferenciação de células-tronco induzida por nanotubos, de forma a nortear o projeto de material e avaliação da biossegurança de implantes modificados por nanotubos.
Materiais e métodos
Fabricação de TiO 2 Nanotubos
Fatias de titânio puro (pureza de 99,9%, espessura de 2 mm; Shengshida, Hebei, China), usadas como substrato, foram polidas com lixa de carboneto de silício de grãos nº 400 e 1500. As amostras foram então lavadas sequencialmente com acetona, álcool anidro e água desionizada em máquina de limpeza ultrassônica e, por fim, secas em temperatura ambiente por 3 h. Para fabricar a nanotopografia, as amostras pré-tratadas foram fixadas como o ânodo, enquanto se usava um pedaço de platina como contra-cátodo em uma solução aquosa de eletrólito de 0,15 M NH 4 F e glicol 90% por 1 h. A voltagem de anodização foi uma voltagem constante de 30, 40, 50, 60 ou 70 V. Após a oxidação anódica, todas as amostras foram enxaguadas com água deionizada por 30 min e lavadas com álcool anidro em máquina de limpeza ultrassônica por 15 min. Por fim, todas as amostras foram esterilizadas em autoclave a 120 ° C por 1 hora e umedecidas com meio de cultura antes do uso.
O mecanismo de reação da fabricação de nanotubos não é claro, e a teoria dominante atual é a teoria de dissolução aprimorada por campo. A formação de arranjos de nanotubos é o resultado do equilíbrio dinâmico sob a ação da oxidação do campo, dissolução do campo e dissolução química (Fig. 1b). O processo de anodização pode ser descrito como segue:na primeira etapa, uma camada de barreira de óxido é formada na interface eletrólito-metal:
$$ {\ mathrm {Ti}} ^ {4 +} + 2 {\ mathrm {H}} _ 2 \ mathrm {O} \ to {\ mathrm {Ti} \ mathrm {O}} _ 2 + 4 {\ mathrm { H}} ^ {+} $$ (1)
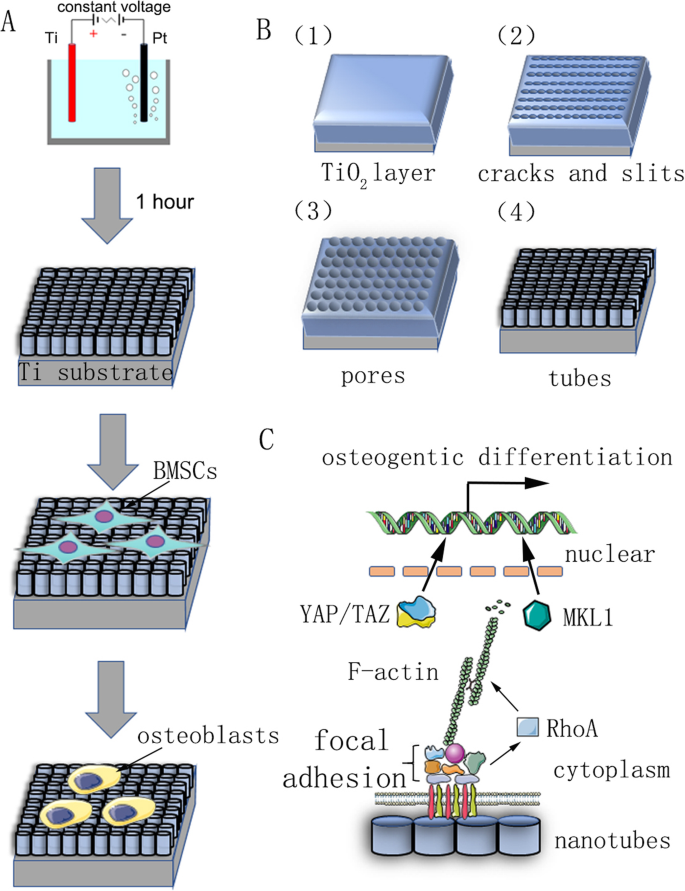
Um gráfico de resumo de nosso estudo. a Fluxograma mostrando a oxidação anódica e a indução da diferenciação celular. b O substrato de titânio pré-tratado foi fixado como o ânodo em uma solução aquosa de eletrólito com 0,15 M NH 4 F e 90% de glicol em voltagem constante por 1 h. Nanotubos automontados foram previstos para serem formados uniformemente. Seu mecanismo de reação é descrito em materiais e métodos. c Diagrama esquemático do mecanismo de diferenciação osteogênica de células-tronco induzidas por nanotubos
Em seguida, rachaduras e fendas estreitas aparecem na superfície devido à dissolução aprimorada pelo campo da camada de óxido. Difusão de F - íons nessas rachaduras e fendas aumentam a taxa de dissolução. As rachaduras aumentam e se conectam com as rachaduras vizinhas. Finalmente, a taxa de formação e a taxa de dissolução da camada de óxido de titânio atingem um equilíbrio dinâmico e os nanotubos não crescem mais:
$$ {\ mathrm {TiO}} _ 2 + 6 {\ mathrm {F}} ^ {-} + 4 {\ mathrm {H}} ^ {+} \ to {\ left [{\ mathrm {TiF}} _ 6 \ right]} ^ {2 -} + 2 {\ mathrm {H}} _ 2 \ mathrm {O} $$ (2)
Caracterização da superfície
Amostras fabricadas com tensões diferentes (30, 40, 50, 60 ou 70 V) foram enxaguadas com etanol e água desionizada por 15 min, depois secas em temperatura ambiente. Microscopia eletrônica de varredura (SEM450, FEI Nova Nano SEM; Thermo Fisher Scientific, Waltham, MA, EUA) foi usada para caracterizar a estrutura da superfície e medir o diâmetro interno e a altura dos nanotubos após o revestimento das amostras com uma fina camada de ouro. Enquanto isso, a análise dispersiva de energia de raios-X (EDS) foi realizada para analisar a composição elementar dos nanotubos. Microscopia de força atômica (AFM, NanoManVS, Bruker Nano Surfaces, Bruker MicroCT, Kontich, Bélgica) foi usada para investigar a morfologia da superfície e a rugosidade da superfície das amostras. Três áreas diferentes foram selecionadas de cada amostra e as medições foram repetidas três vezes.
Cultura de células
Ratos Sprague-Dawley (SD) machos de quatro semanas de idade foram adquiridos no centro experimental de animais do Shanghai Ninth People’s Hospital (Shanghai, China). Células-tronco mesenquimais da medula óssea de rato (BMSCs) foram isoladas assepticamente de fêmures e tíbias. As BMSCs foram purificadas e expandidas em meio essencial mínimo α (α-MEM; Hyclone, Logan, UT, EUA) contendo 10% (v / v) de soro fetal bovino (FBS) (Gibco / Life Technologies, Carlsbad, CA, EUA ), Estreptomicina 100 mg / mL (Gibco) e penicilina 100 U / mL (Gibco), e incubados a 37 ° C em uma atmosfera umidificada consistindo em 95% de ar e 5% de CO 2 . O meio de cultura foi renovado a cada 2 dias, e as células foram tripsinizadas e subcultivadas a 80% de confluência. Todas as células usadas neste estudo estavam entre as passagens 3 e 5. O meio de indução osteogênico era composto de meio de crescimento suplementado com dexametasona 100 nM, β-glicerofosfato 10 mM e ácido ascórbico 50 mM (Sigma-Aldrich, St Louis, MO, EUA) .
Ensaio de proliferação celular
O TiO 2 fatias de titânio modificadas com nanotubos foram cortadas em formas circulares e colocadas em poços de uma placa de cultura de células de 24 poços. BMSCs entre as passagens 3-5 foram cultivadas em TiO 2 nanotubos com densidade de 3 × 10 4 células / disco em meio de crescimento ou meio osteogênico. Após 2 dias de cultura de células, citocalasina D (Cyto D, Sigma-Aldrich) e jasplakinolide (Jasp, Sigma-Aldrich), usados para interromper a polimerização de F-actina, foram adicionados ao meio todos os dias por 3 dias. A concentração final e o tempo de trabalho de Cyto D e Jasp foram 5 μM, 1 he 2 μM, 3 h, respectivamente. O meio de cultura foi renovado após incubação com os reagentes. A viabilidade e proliferação celular foram avaliadas usando o ensaio Cell Counting Kit-8 (CCK8) (Dojindo, Kumamoto, Japão) 12 h após o tratamento com Cyto D ou Jasp. As células foram incubadas com solução de CCK8 a 10% (v / v) por 2 h em uma incubadora de células a 37 ° C sob 5% de CO 2 . Em seguida, transferimos 100 μL da mistura de reação para poços de uma placa de 96 poços e a absorbância (DO) do produto corante de formazan na cultura foi medida a 450 nm usando um espectrofotômetro Multiscan UV-visível (Safire2; TECAN, Mannedorf, Suíça ) Além disso, também contamos o número aproximado de células usando um contador automatizado de células (AMQAX1000, Life Technologies). Antes da contagem das células, os BMSCs foram enzimaticamente separados do TiO 2 nanotubos e corados com azul Trypan (Sigma – Aldrich).
Coloração com Fosfatase Alcalina e Análise da Atividade ALP
BMSCs foram semeados em cinco TiO 2 diferentes fatias de titânio modificadas por nanotubos (30, 40, 50, 60 ou 70 V) a uma densidade de 3 × 10 4 por poço e cultivadas em meio osteogênico. Os reagentes foram adicionados conforme descrito acima. Após 7 dias de incubação, BMSCs cultivados em TiO 2 nanotubos foram lavados três vezes com PBS, fixados com paraformaldeído 4% e incubados em solução de trabalho de fosfatase alcalina (ALP) de um kit ALP de acordo com as instruções do fabricante (Hongqiao, Shanghai, China). Os resultados foram observados em um estereomicroscópio após lavagem com PBS.
Para a análise da atividade de ALP, as células foram primeiro lisadas com tampão RIPA sem inibidores de protease e fosfatase e, em seguida, os lisados centrifugados foram testados usando um kit de ensaio ALP (Beyotime Institute of Biotechnology, Jiangsu, China) seguindo o protocolo fornecido. A atividade foi finalmente normalizada para a concentração de proteína do lisado correspondente.
Imunocitoquímica
Após 3 dias de tratamento com Cyto D e Jasp, as BMSCs foram fixadas com paraformaldeído a 4% por 20 min em temperatura ambiente e depois lavadas três vezes com PBS. As células foram permeabilizadas com 0,3% de Triton-X 100 por 30 min, lavadas três vezes com PBS e coradas com faloidina conjugada a rodamina por 1 h em temperatura ambiente no escuro. Em seguida, as células foram enxaguadas com PBS e contrastadas com DAPI (Beyotime Institute of Biotechnology) por 10 min em temperatura ambiente. Após três lavagens adicionais com PBS, as amostras foram fixadas em uma lâmina de vidro e observadas por microscopia confocal.
Western Blotting
Para avaliar a expressão da proteína, BMSCs cultivados em TiO 2 nanotubos foram colhidos com tripsina (Gibco). As células foram lavadas três vezes com PBS e lisadas com tampão RIPA suplementado com um coquetel de inibidor de protease e fosfatase por 30 min em gelo. O lisado foi coletado por centrifugação a 12.000 × g por 15 min a 4 ° C. A concentração de proteínas totais no sobrenadante foi medida usando um kit de ensaio de proteína de ácido bicinconínico (BCA) (Beyotime) de acordo com as instruções do fabricante. O tampão de carregamento foi adicionado às amostras de proteína como acima, que foram então fervidas a 95 ° C durante 15 min. Para a análise de Western blotting, 10 μL da preparação de proteína foram carregados em um gel de SDS-PAGE 12,5% (EpiZyme Inc., Cambridge, MA, EUA) e submetidos a eletroforese a 120 V por 1 h, em seguida, eletrotransferidos para um polivinilidenodifluoreto (PVDF) membrana a 250 mA durante 2 h. As membranas foram então bloqueadas com 5–10% de leite em pó desnatado em TBST por 1 h em um agitador em temperatura ambiente e incubadas com anticorpo primário diluído em tampão de diluição (Beyotime) a 4 ° C durante a noite. Em seguida, anticorpo secundário conjugado com fluorescência diluído em tampão de diluição foi adicionado às membranas após lavagem três vezes com TBST por 5 min, e as membranas foram então incubadas em temperatura ambiente por 1 h no escuro. As bandas de proteína foram detectadas por um sistema de imagem de fluorescência infravermelha de duas cores (Odyssey, LiCor Biosciences, Lincoln, NE, EUA). Em particular, se as bandas da proteína de referência interna foram unificadas, a membrana foi removida e reexaminada com outro anticorpo primário, seguido pelo mesmo processo. Usamos o anticorpo GAPDH como nossa proteína de referência interna para normalizar a expressão da proteína, e os outros anticorpos primários usados neste estudo foram anti-vinculina (diluição 1:1000, Abcam, Cambridge, MA, EUA), anti-FAK (diluição 1:1000 , Cell Signaling Technology, Danvers, MA, EUA), anti-Runx2 (diluição 1:1000, Cell Signaling Technology), anti-RhoA (diluição 1:1000, Cell Signaling Technology), anti-F-actina (diluição 1:500 , Abcam), anti-Osx (diluição 1:500, Abcam) e anti-pYAP (diluição 1:1000, Cell Signaling Technology). Os anticorpos secundários foram IgG H&L de cabra anti-rato (IRDye® 680RD, diluição 1:5000, Abcam) e IgG H&L de cabra anti-coelho (IRDye® 680RD, diluição 1:5000, Abcam).
PCR quantitativo em tempo real
PCR quantitativo em tempo real foi realizado no dia 7 para avaliar a expressão gênica do fator de transcrição relacionado a runt 2 (Runx2), Osterix (Osx), Alp, osteocalcina (OCN), RhoA, YAP, TAZ, vinculina (VCL), cinase de adesão focal (FAK) e leucemia megacarioblástica 1 (MKL1) em células cultivadas em meio osteogênico em TiO 2 nanotubos. O RNA total foi extraído das células usando um kit de RNA total (R6812-01HP, Omega Bio-Tek Inc., Norcross, GA, EUA). A concentração e a pureza das amostras de RNA foram determinadas por densidade óptica em um comprimento de onda de 260 e apenas as amostras apresentando razões A 260/280 e A 260/230 maiores que 1,8 foram analisadas. As amostras de RNA foram transcritas reversamente em cDNA usando um kit qScript cDNA Synthesis (Takara, Shiga, Japão) de acordo com as instruções do fabricante. A PCR quantitativa em tempo real foi realizada com SYBR® Premix Ex Taq ™ (Takara) usando um sistema de PCR em tempo real QuantStudio 6 Flex (Life Technologies). GAPDH, um gene de manutenção da casa, foi usado como uma referência interna. Os dados foram analisados usando a comparação Ct (2 -ΔΔCt ) método e expresso como alterações em dobras em comparação com o controle. As sequências dos primers usados estão listadas na Tabela 1.
Análise estatística
Todos os dados são representativos de pelo menos três experiências independentes usando amostras em triplicado, salvo indicação em contrário. Os dados são expressos como média ± desvio padrão (DP). As diferenças entre os grupos foram avaliadas por análise de variância unilateral seguida pelo teste post-hoc de Student-Newman-Keuls ou t de Student teste. P valores <0,05 foram considerados estatisticamente significativos.
Resultados
Caracterização da superfície
Para fabricar a nanotopografia, TiO 2 nanotubos foram formados em um substrato de titânio puro usando equipamento de oxidação anódica em diferentes tensões constantes (30, 40, 50, 60 e 70 V) por 1 h (Fig. 1). Uma matriz uniformemente distribuída de nanotubos automontados foi observada por microscopia eletrônica de varredura (SEM). As vistas lateral e superior dos nanotubos são mostradas na Fig. 2a, b. A altura dos nanotubos em todas as amostras neste estudo foi de aproximadamente 2 μm, enquanto os diâmetros internos dos nanotubos foram de aproximadamente 74 nm (30 V), 92 nm (40 V), 112 nm (50 V), 128 nm (60 V) e 148 nm (70 V) (Fig. 2c). Isso indicou que a altura dos nanotubos estava relacionada ao tempo de oxidação anódica e o diâmetro interno estava relacionado à voltagem de oxidação anódica. A análise dispersiva de energia de raios-X (EDS) foi então realizada para analisar a composição elementar dos nanotubos. Isso mostrou que os nanotubos consistem em apenas dois elementos, O e Ti (Fig. 2d, e). A microscopia de força atômica (AFM) foi utilizada para detectar as estruturas nanotubulares e medir o desvio médio aritmético do perfil dos nanotubos conforme a rugosidade da superfície (Ra) (Fig. 2f). Os dados mostraram que a rugosidade da superfície dos nanotubos aumentou à medida que o diâmetro (ou seja, a voltagem de oxidação anódica) aumentou (Fig. 2g).
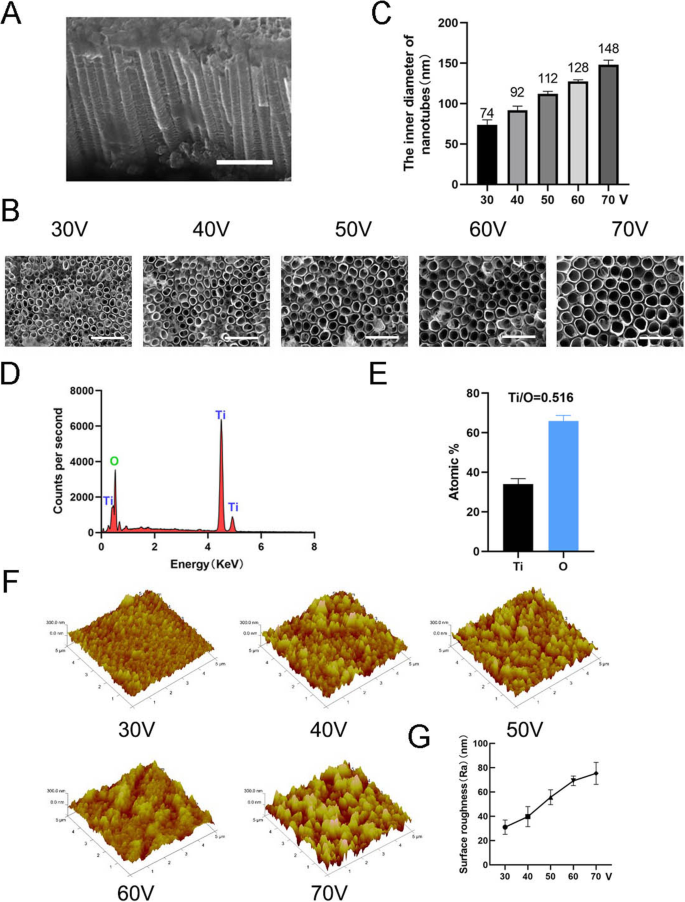
Caracterização de superfícies de nanotubos. a Vista lateral dos nanotubos. Barras de escala:1 μm. b Vista superior dos nanotubos. Barras de escala:500 nm. c O diâmetro interno dos nanotubos em cinco tensões constantes diferentes (30, 40, 50, 60, 70 V). d A composição do elemento químico EDS de estruturas de nanotubos. e A proporção do elemento de Ti e O. f Imagens de microscopia de força atômica (AFM) de estruturas de nanotubos. g A rugosidade superficial média (Ra) dos nanotubos
TiO 2 Diferenciação Osteogênica Induzida por Nanotubos de MSCs
Após 7 dias de osteoindução, a coloração de ALP foi primeiro realizada para avaliar a diferenciação osteogênica de MSCs. Os resultados da coloração mostraram que os MSCs cultivados em TiO 2 nanotubos tiveram maior atividade ALP do que células cultivadas em um substrato de titânio liso (grupo de controle) (Fig. 3a). A análise estatística da área de coloração demonstrou que a capacidade dos nanotubos de induzir a diferenciação osteogênica foi significativamente aumentada quando comparada ao grupo controle. Enquanto isso, observamos uma tendência de que dentro da faixa de diâmetro deste experimento, maior será o diâmetro do TiO 2 nanotubos, maior será a capacidade de induzir a diferenciação osteogênica (Fig. 3b). Assim, o grupo de 70 V foi usado em experimentos subsequentes para melhor exibir os resultados. Em seguida, analisamos a expressão do gene osteogênico nos dias 3 e 7. MSCs cultivadas em TiO 2 nanotubos por 3 dias e 7 dias, ambos mostraram promoção significativa da expressão de genes osteogênicos (RUNX2, ALP, OCN e OSX) em comparação com o grupo de controle (Fig. 3d-g). Os resultados do Western blot confirmaram que a expressão da proteína de RUNX2 e OSX também aumentou após 7 dias de osteoindução (Fig. 3c). Curiosamente, descobrimos que a F-actina foi regulada positivamente no TiO 2 grupo de nanotubos. Portanto, era óbvio que TiO 2 nanotubos direcionaram MSCs para a diferenciação de osteoblastos, que estava relacionada ao diâmetro dos nanotubos. Nossos resultados também sugeriram o envolvimento da F-actina neste processo.
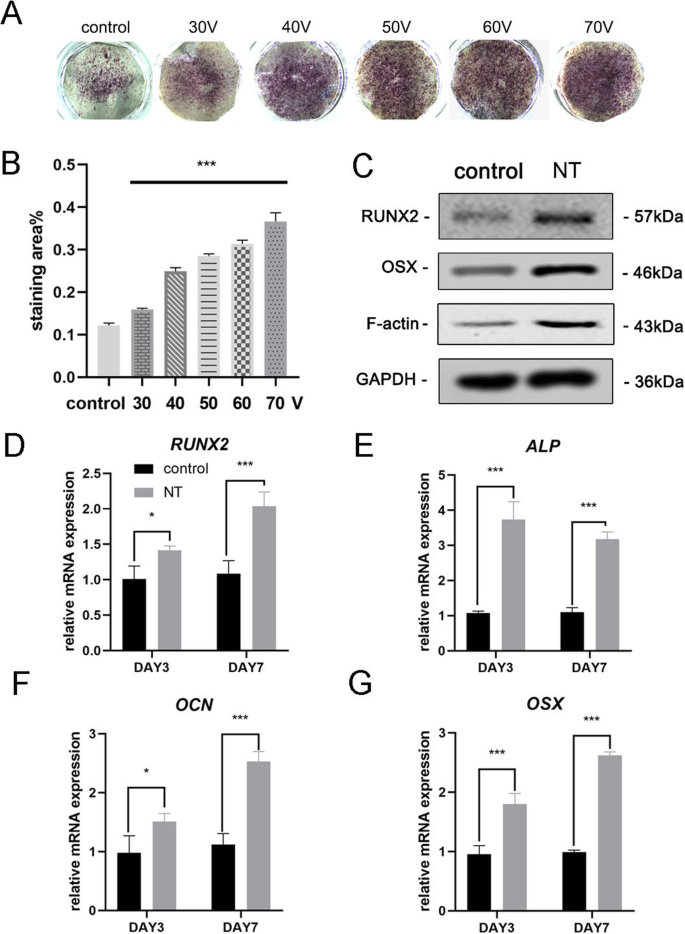
TiO 2 nanotubos aumentaram a expressão do gene osteogênico de BMSCs. a Coloração ALP do substrato de titânio liso e cinco substratos de nanotubos diferentes. As células foram induzidas com meio osteogênico por 7 dias. b A análise estatística da área de coloração foi realizada usando ImageJ. c Proteínas associadas à osteogênese (RUNX2 e OSX) e F-actina em MSCs foram analisadas por Western blotting no dia 7. A expressão de mRNA de RUNX2 ( d ), ALP ( e ), OCN ( f ) e OSX ( g ) nos dias 3 e 7, analisados por qRT-PCR. NT o grupo de nanotubos. Os dados representam a média ± DP de três amostras. * P <0,05, ** P <0,01 e *** P <0,001
Diferenciação de osteoblasto mediada por F-actina de MSCs em TiO 2 Nanotubos
Para explorar ainda mais se F-actina estava envolvida na diferenciação celular induzida por nanotopografia, usamos dois reagentes, jasplakinolide (Jasp) e citocalasina D (Cyto D), para regular a polimerização de F-actina de forma positiva e negativa, respectivamente. As fotomicrografias confocais de coloração com rodamina-faloidina mostraram que F-actina no grupo de tratamento Cyto D foi quase despolimerizada e estruturas fibrosas foram raramente vistas, enquanto Jasp estabilizou e polimerizou F-actina, verificada por estruturas semelhantes a feixes mais distintas e brilhantes do que as observadas no grupo controle (Fig. 5a). Além disso, a análise de Western blot também confirmou que a expressão da proteína F-actina foi afetada, o que provou que Cyto D e Jasp agiram como esperado (Fig. 5b). O ensaio de proliferação celular mostrou que Cyto D inibiu significativamente a proliferação celular, enquanto Jasp promoveu o crescimento celular (Fig. 4a). Os resultados da contagem de células foram consistentes com este achado (Fig. 4b).
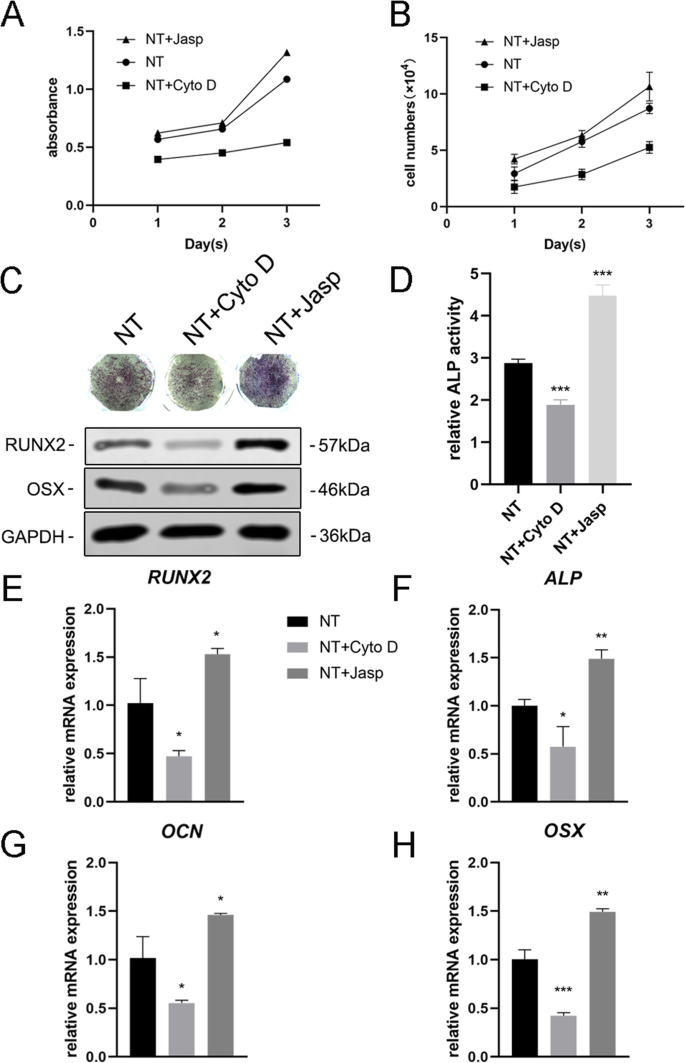
A montagem da F-actina regulou a expressão de genes osteogênicos em BMSCs. a , b A proliferação celular após o tratamento com Cyto D e Jasp foi determinada usando o ensaio CCK-8 ou contador automatizado de células nos dias 1, 2 e 3. c , d A coloração de ALP e a análise da atividade de ALP foram realizadas para avaliar a expressão de ALP em MSCs após o tratamento com drogas por 3 dias. A área de coloração foi analisada usando ImageJ. c , e - h Western blotting e qRT-PCR foram usados para comparar as alterações dos marcadores relacionados à osteogênese no grupo NT + Cyto D e no grupo NT + Jasp com aqueles do grupo controle (sem tratamento medicamentoso). NT o grupo dos nanotubos. Os dados representam a média ± DP de três amostras. * P <0,05, ** P <0,01 e *** P <0,001
Em seguida, avaliamos a capacidade das MSCs de se diferenciarem em osteoblastos para investigar se a F-actina mediava esse processo. Primeiro, detectamos ALP como um marcador precoce de osteogênese. Em comparação com o grupo de controle, o tratamento com Cyto D reduziu a expressão de ALP e sua atividade, enquanto que no grupo de tratamento com Jasp foi regulado positivamente (Fig. 4c, d). Consistente com este resultado, o tratamento com Jasp resultou em um aumento nos níveis de proteína de RUNX2 e OSX, enquanto Cyto D teve o efeito oposto (Fig. 4c). Em concordância com isso, os níveis de expressão de mRNA de genes osteo-específicos, incluindo RUNX2, ALP, OCN e OSX, mostraram a mesma tendência após o tratamento medicamentoso (Fig. 4e-h). Acima de tudo, esses dados indicaram que F-actina desempenhou um papel importante no processo de diferenciação osteogênica de MSCs induzida por TiO 2 nanotubos. A promoção da despolimerização da F-actina inibiu a diferenciação de osteoblastos induzida por nanotopografia, enquanto a estabilização e a polimerização da F-actina aumentaram a diferenciação de osteoblastos.
Diferenciação de Osteoblastos Regulados por F-actina de MSCs em TiO 2 Nanotubos através de MKL1 e YAP / TAZ
Para dissecar o mecanismo subjacente que envolve a F-actina na regulação do destino das MSCs, investigamos proteínas / moléculas que interagem diretamente com a F-actina ou afetam a polimerização da F-actina. Primeiro, tentamos identificar como a nanotopografia afetou o equilíbrio entre F-actina e G-actina. TiO 2 nanotubos como um sinal físico diferem dos sinais químicos permeáveis à membrana e, portanto, devem usar alguns componentes da membrana celular para transmitir estímulos para as células. O acúmulo de evidências indica que o complexo de adesão focal, incluindo integrina, talina, cinase de adesão focal (FAK), vinculina (VCL), tensina e outras proteínas, funciona como um portador de sinal, que informa as células sobre a condição da matriz extracelular (ECM ) e, portanto, afeta seu comportamento biológico [22, 23]. Mais importante, a F-actina se liga às integrinas por meio de tais complexos de adesão focal e, assim, forma ligações mecânicas entre os feixes de actina intracelular e a ECM [24]. Consequentemente, analisamos a seguir a expressão dos componentes do complexo de adesão focal. Os resultados mostraram que a expressão da proteína e do mRNA de VCL e FAK foram consistentes com a alteração da F-actina, indicando que o complexo de adesão focal estava envolvido no processo de diferenciação osteogênica de MSCs induzida por TiO 2 nanotubos (Figs. 5b e 6a, b). Além disso, também descobrimos que RhoA, uma pequena proteína GTPase na família Rho de GTPases, foi regulada positivamente no grupo de tratamento Jasp e inibida por Cyto D (Figs. 5b e 6a, b). RhoA é uma importante molécula de transdução de sinal a montante na via MAPK e pode ser regulada por FAK [25, 26]. A função primária do RhoA é promover a polimerização e estabilidade das fibras de estresse (F-actina) e montagem do complexo de adesão focal [27]. Juntos, esses dados sugerem que TiO 2 nanotubos podem influenciar a polimerização de F-actina por meio do complexo de adesão focal e RhoA.
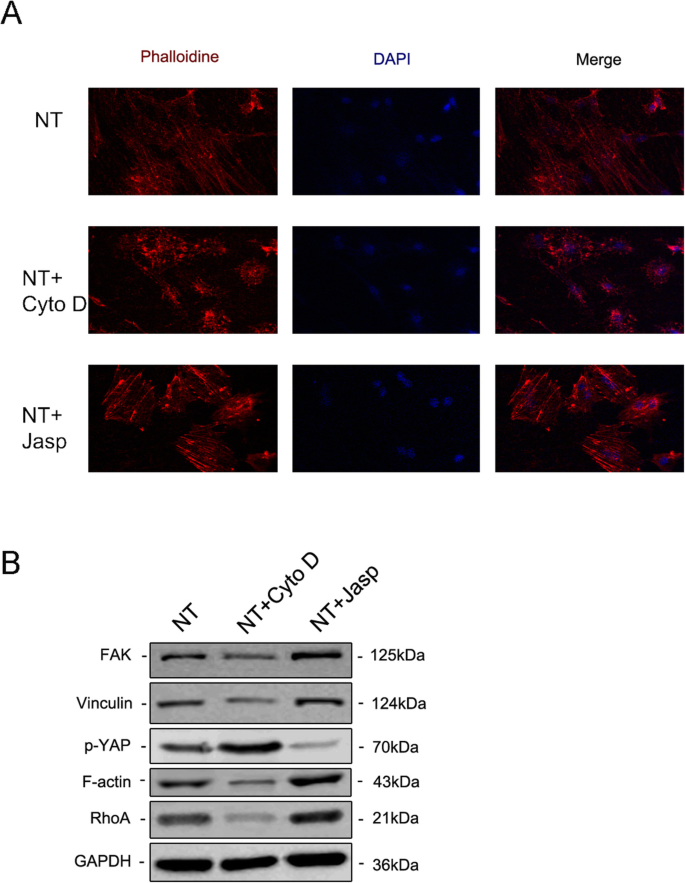
A coloração de imunofluorescência revelou o nível de F-actina pela coloração com faloidina conjugada com rodamina ( a ) A expressão da proteína de FAK e VCL contida no complexo de adesão focal, RhoA e YAP fosforilado foram investigadas por Western blotting ( b ) NT o grupo de nanotubos

Effect of F-actin assembly on FAK (a ), vinculin (VCL) (b ), RhoA (c ), MKL1 (d ), YAP (e ), and TAZ (f ) gene expression in MSCs. NT the nanotubes group. Data represent the mean ± SD of three samples. * P <0.05, **P <0.01, and ***P <0,001
But how does F-actin regulate cell fate? Most studies have demonstrated that F-actin is involved in cell migration, cell division, endocytosis, and especially tumor cell invasion [28,29,30]. Few studies have suggested that F-actin could also regulate cell differentiation, let alone its specific molecular mechanism [31, 32]. Consequently, we searched for articles that mentioned the F-actin changes and found that YAP/TAZ, two closely related transcriptional co-activators in the Hippo signaling pathway, which shuttle between the cytoplasm and the nucleus, may serve as mechanotransducers in regulating MSC differentiation [33,34,35]. In addition, we also found that MKL1, a key regulator of smooth muscle cell differentiation, which interacts with the transcription factor serum response factor, could bind to G-actin and also circulate between the cytoplasm and the nucleus [21, 36]. Our results ultimately identified the involvement of YAP/TAZ and MKL1 in nanotube-induced osteoblast differentiation mediated by F-actin (Figs. 5b and 6d–f). Interestingly, the protein expression of phosphorylated YAP showed the opposite trend, indicating that not only was the expression of YAP changed, but the phosphorylation of YAP was also changed by Cyto D and Jasp (Fig. 5b). This result was consistent with the report that the phosphorylation of YAP/TAZ could be sequestrated in the cytoplasm [35].
In summary, our results preliminarily demonstrated that F-actin regulated osteoblast differentiation of MSCs on TiO2 nanotubes through MKL1 and YAP/TAZ (Fig. 7).

Schematic representation of F-actin assembly induced by nanotubes, and the putative role of MKL1 and YAP/TAZ in acting as the downstream mediators of F-actin signaling to regulate gene expression
Discussão
Titanium and titanium alloys are the most widely used metal materials in orthopedic clinical implants due to the good properties of titanium [3]. However, aseptic loosening is still an urgent problem to be solved and improved, and the key is likely to lie in improving the integration of the implant and the host bone. Previous studies have shown that surface coating and modification or immobilization of biofunctional molecules will be beneficial to osseointegration [37]. Recently, the surface topography of implants has attracted the attention of many researchers thanks to studies into the cell response to physical cues [9,10,11, 13, 38]. In this study, we demonstrated the ability of nanotubes to promote osteogenic differentiation of MSCs, and this ability was enhanced with increasing inner diameter of the nanotubes (30–70 V). This will help guide the diameter of the nanotubes on the surface of the implants.
As a topographical structure, nanotubes first change the physical properties of the material, such as adsorption capacity and electrical and thermal conductivity. These physical properties determine their application in the industrial field. For example, most high-voltage power transformers need to be filled with insulating material, which is usually transformer oil or insulating gas. When the insulation of a transformer fails due to overheating and partial discharge, a serious discharge accident will occur. Therefore, finding an effective method that accurately detects the concentration and types of dissolved gases or insulating gas decomposition components in a transformer is necessary to monitor the operating state of the transformer [8, 39,40,41]. The traditional approach is to look for materials with good gas adsorption in transition elements, which are rich in d electrons, such as Pd(1 1 1) [39]. Nowadays, nanotubes are widely studied for their good gas adsorption properties. He et al. found that CuO-BNNT was suitable for the adsorption of C2 H 2 , because of its stronger adsorption on C2 H 2 [8]. Meanwhile, TiO2 itself can be a gas-sensing material. Gui et al. found that Co-doped TiO2 further enhanced gas adsorption capacity and exhibited a superior adsorption ability and conductivity change toward C2 H 4 molecules [40]. Consistent with this study, Mn-doped graphene also exhibited enhanced conductivity and superior capability of C2 H 2 and CO detection than pristine graphene [41]. The above research indicates that the TiO2 nanotubes prepared in our experiment have a potential application in the field of monitoring the operative state of a transformer. However, the adsorption capacity and electrical conductivity of the nanotubes to gases need to be further studied, especially whether these properties are enhanced after doping with transition elements (e.g., Mn).
In addition to changing the physical properties of a surface, nanoscale morphology also affects the biological behavior of the cells attached to it. Cells first adhere to the surface of the material and then migrate, proliferate, and differentiate. Compared to a flat surface, the hollow structure of the nanotubes provides fewer adhesion sites for cells. Therefore, in order for the cells to adhere to the nanotube surface steadily and maintain the biomechanical balance within the cell, the focal adhesion complex begins to assemble and mature, and F-actin becomes strong and stable.
F-actin, a linear polymer microfilament consisting of G-actin monomers, is one of the three major components of the cytoskeleton. As a mechanical-loading structure, F-actin is generally believed to be involved in cell division, cell migration, endocytosis, and tumor cell invasion [28,29,30], but some recent studies showed that it can also affect cell differentiation [31,32,33, 36]. For example, actin cytoskeletal depolymerization by simvastatin induces chondrocyte differentiation [31], and actin depolymerization enhances adipogenic differentiation in human stromal stem cells [32]. Our results also revealed that, compared with the control group, MSCs cultured on nanotubes had higher F-actin levels and a more obvious fibrous structure. Meanwhile, promotion of F-actin polymerization by Jasp enhanced osteogenic differentiation, while the depolymerization of F-actin inhibited osteogenic differentiation, suggesting that F-actin mediates TiO2 nanotube-induced osteoblastic differentiation of MSCs.
F-actin can be regulated by Rho GTPases, members of the Ras superfamily [23, 42], and Rho can induce actin reorganization through at least two effectors, ROCK and Dia. ROCK is activated by binding to Rho-GTP and then myosin light chain (MLC), the substrate of ROCK, plays an important role in F-actin assembly. ROCK inhibits the activity of MLC phosphatase, leading to an increase in MLC phosphorylation, which stimulates the ATPase activity of myosin II and promotes the assembly of F-actin. In addition, ROCK also targets LIM kinase (LIMK). Phosphorylated LIMK inactivates cofilin by phosphorylation, which can disassemble F-actin in its active state. Another effector is Dia, a member of the formin-homology (FH) family of proteins which contains two FH domains. These domains contain multiple proline-rich motifs which bind to the G-actin-binding protein, profilin. This interaction contributes to actin polymerization and F-actin organization [42]. We detected one of the Rho GTPases, RhoA, and found that the expression of RhoA was consistent with the level of F-actin. However, we were unable to clearly describe how the nanotubes regulate the expression of RhoA, because there are many other regulators, including integrin signaling, other adhesion receptors, G protein-coupled receptors (GPCRs), soluble factors such as LPA, receptor tyrosine kinase signaling, and so on [43].
Knowing that F-actin can be regulated by RhoA, we next asked what role focal adhesion played in this process, because focal adhesion complexes, containing integrins, talin, vinculin, paxillin, and focal adhesion kinase (FAK), are formed and mature when cells attach to the surface of nanotubes. Integrins are transmembrane heterodimers that couple the ECM to the other focal adhesion proteins so as to facilitate cell attachment. They not only act simply as hooks but also transmit to the cell critical signals about the nature of its surroundings, which along with other signals such as EGFR, prompt the cell to make decisions about its biological behaviors. These signals are further transmitted to F-actin, which is directly connected to the focal adhesion complexes. On the one hand, the nanoscale morphology causes focal adhesion complex assembly and maturation. On the other hand, kinases such as FAK and Src kinase family members will recruit molecules such as CRK to self-regulate the assembly and maturation of focal adhesion complexes [44,45,46]. Our results demonstrated that the formation and maturation of focal adhesion complexes were impaired by F-actin depolymerization, suggesting that there was a feedback from focal adhesion complexes to actin assembly in line with published reports.
However, it should not be ignored that these proteins contained in focal adhesion complexes have the function of signal transduction [47]. That is to say, nanotubes may directly regulate gene expression through signal cascades, and F-actin may just participate in or be affected by this process. For instance, the dual kinase complex of FAK and Src can regulate Rho GTPases such as RhoA. This shows that nanotubes can regulate RhoA through integrins and the FAK/Src complex. In addition Src, a non-receptor tyrosine kinase protein, can activate Ras (small GTPase) by phosphorylating FAK at tyrosine residue 925 [47, 48]. Then, Ras activates numerous biochemical pathways, including the well-studied MAPK pathway and the PI3K/AKT/mTOR pathway. In the MAPK pathway, Ras activates c-Raf, followed by mitogen-activated protein kinase kinase (MAP2K) and then MAPK1/2, also known as extracellular signal-regulated kinase (ERK). ERK in turn activates transcription factors such as serum response factor (SRF) and c-Myc that are involved in regulating growth and differentiation [49]. What is more, Runx2, a key transcription factor in osteogenic differentiation, can also be regulated by ERK [50], and our previous study confirmed that mechanical strain promoted osteogenic differentiation of BMSCs through the FAK-Erk1/2-Runx2 pathway [17]. Therefore, we cannot rule out that ERK plays a role in nanotube-induced osteogenic differentiation and further study is still needed.
So what exactly is the role of F-actin in inducing differentiation of nanotubes, because its change can affect cell differentiation? One possibility is that the change of F-actin assembly can inversely regulate the level of FAK so as to induce osteogenic differentiation through the FAK-Erk1/2-Runx2 pathway as described above, because in our results, focal adhesion complexes and actin polymerization showed the same trends of change, indicating that they act as a whole in response to the extracellular environment. However, some other possibilities also exist, and a number of articles have shown that MKL1 and YAP/TAZ act downstream of the actin dynamic balance [20, 51,52,53,54]. Both of them shuttle between the cytoplasm and the nucleus, and may help to transduce signals from the cytoskeleton to the nucleus.
MKL1, also termed myocardin-related transcription factor A, is sensitive to changes in G-actin levels. When cytoplasmic G-actin levels increase, monomeric G-actin binds to MKL1 and prevents it from binding to SRF and activating transcription. SRF target genes include actins such as smooth muscle actin (SMA) as well as other actin-binding proteins, including immediate early genes like c-fos and egr1. Recent studies have demonstrated that changing SRF activity could regulate adipogenesis by activating the adipogenesis transcription factor peroxisome proliferator-activated receptor γ (PPARγ), and also regulate bone formation via IGF-1 and Runx2 signaling [55, 56].
YAP and TAZ are two transcriptional coactivators in the Hippo signaling pathway, identified as an important regulatory pathway that restricts cell proliferation, thereby controlling organ size and morphogenesis [20]. Large tumor suppressor genes 1 and 2 (LATS1/2) phosphorylate them, thereby creating a binding site for 14-3-3 proteins, the binding of which prevents their nuclear import [53, 54]. As a consequence, phosphorylated forms of YAP/TAZ are sequestered in the cytoplasm, preventing the expression of genes like Ctgf and Areg. In addition, some studies have shown that YAP/TAZ can interact with T-box 5 (TBX5), RUNX2, and p73 to regulate gene expression [57,58,59]. Further, cell adhesion to cell matrix proteins has been shown to trigger YAP nuclear localization through an integrin/FAK/Src axis. In our study, the results suggested that this pathway was possibly involved in nanotube-induced differentiation. Further study into the downstream mediators of the integrin/FAK/Src axis should be carried out to clarify the specific mechanism.
On the other hand, more and more studies illustrate that F-actin interacts with Hippo signaling, and somehow inhibits the phosphorylation of YAP [54, 60], which is consistent with our experimental results that promoting F-actin polymerization reduces the expression of phosphorylated YAP. We hypothesize that ATP involved in the process of the transformation between G-actin and F-actin may also play an important role in the phosphorylation of YAP, which is yet to be studied.
After understanding the above possible molecular mechanisms, we can try to explain some of the experimental phenomena found in this study. Our results revealed that the larger the diameter of the nanotubes, the stronger the ability of the nanotubes to promote osteogenic differentiation. This is consistent with previous research [61, 62]. The reason for this phenomenon is that the larger the diameter of the nanotubes, the less adhesion sites they can provide to the cells, and the greater the assembly and maturity of focal adhesion complexes. Along with these, stress fibers made of F-actin will have greater strength and stability. These structures enhance the signaling that promotes osteogenic differentiation. Predictably, however, this effect is significantly reduced when the nanotubes become too large in diameter, making it difficult for the cells to adhere to the surface [12]. Similarly, when the height of the nanotubes is inconsistent, the differences in height can result in a change of adhesion site and rearrangement of the cytoskeleton, which will further affect cell differentiation. Intriguingly, even flat surface materials without nanotube modification can induce changes in cell differentiation. A number of studies have demonstrated that focal adhesion formation and stress fiber organization are regulated by substrate stiffness [63,64,65], and YAP/TAZ also plays an important role in this process. Therefore, it is obvious that the integrins–FAs (focal adhesions)–F-actin axis plays a role in the transduction of physical signals into intracellular chemical signals.
In summary, our results demonstrated that F-actin regulates osteoblastic differentiation of mesenchymal stem cells on TiO2 nanotubes through MKL1 and YAP/TAZ, whose target genes partly explained the proliferation and differentiation of MSCs. We know that there is no single change in the signal network and any change is regulated by numerous molecules and proteins. One type of biological behavior must be the result of the regulation of a series of signaling pathways. Nanotubes induce cell differentiation by triggering a complex network of signals, including integrins, proteins contained in focal adhesion complexes, FAK, Src, Rho GTPase, the MAPK pathway, the Hippo pathway, and other reported signaling pathways. At least as important, there are many signal cycles in the signal network and a downstream signal can regulate the upstream signal via feedback. In this study, we found that vinculin and FAK can be regulated backwards by F-actin assembly, increasing the uncertainty of molecular function. Therefore, more details of the molecular mechanism await further study.
Conclusões
Our results showed that TiO2 nanotubes promoted the osteogenic differentiation of MSCs, and this ability was enhanced with the increasing diameter of nanotubes within a certain range (30–70 V). F-actin mediated nanotube-induced cell differentiation through MKL1 and YAP/TAZ, providing a novel insight into the study of cell differentiation.
Disponibilidade de dados e materiais
The datasets used and analyzed during the current study are available from the corresponding author on reasonable request.
Abreviações
- MSCs:
-
Células-tronco mesenquimais
- SEM:
-
Microscopia eletrônica de varredura
- EDS:
-
X-ray energy dispersive analysis
- AFM:
-
Força atômica microscópica
- ALP:
-
Alkaline phosphatase
- Cyto D:
-
Cytochalasin D
- Jasp:
-
Jasplakinolide
- VCL:
-
Vinculin
- FAK:
-
Focal adhesion kinase
- BCA:
-
Bicinchoninic acid
- PVDF:
-
Polyvinylidenedifluoride
- Runx2:
-
Runt-related transcription factor 2
- Osx:
-
Osterix
- OCN:
-
Osteocalcin
- YAP:
-
Yes-associated protein
- MKL1:
-
Megakaryoblastic leukemia 1
- FBS:
-
Soro fetal bovino
- ECM:
-
Extracellular matrix
- MLC:
-
Myosin light chain
- LIMK:
-
LIM kinase
- FH:
-
Formin-homology
- GPCR:
-
G protein-coupled receptors
- MAP2K:
-
Mitogen-activated protein kinase kinase
- ERK:
-
Extracellular signal-regulated kinase
- SRF:
-
Serum response factor
- SMA:
-
Smooth muscle actin
- PPARγ:
-
Peroxisome proliferator-activated receptor γ
- LATS1/2:
-
Large tumor suppressor gene 1 and 2
- TBX5:
-
T-box 5
Melhorando as propriedades eletroquímicas de LaCoO3 por Sr-Doping, rGO-Compounding com Rational Design for Energy Storage Device
Pontos de carbono como materiais de nova geração para nanotermômetro:Revisão
Nanomateriais
- DIÓXIDO DE TITÂNIO - TiO2 - Preço, mercado e análise
- Em direção aos nanofluidos de TiO2 - Parte 1:Preparação e propriedades
- Nanotécnicas Inativam células-tronco cancerosas
- Rumo aos nanofluidos de TiO2 - Parte 2:Aplicações e desafios
- Efeito da distribuição de nanopartículas de ouro em TiO2 nas características ópticas e elétricas de células solares sensibilizadas por corante
- Síntese de nanocristais de ZnO e aplicação em células solares de polímero invertido
- O precursor de titânio ideal para a fabricação de camada compacta de TiO2 para células solares de perovskita
- Influência da rigidez elástica e adesão da superfície no salto de nanopartículas
- Influência de nanopartículas de Ag com diferentes tamanhos e concentrações incorporadas em uma camada compacta de TiO2 na eficiência de conversão de células solares de perovskita
- Um novo material de conversão ascendente de Ho3 + -Yb3 + -Mg2 + TiO2 tri-dopado e suas aplicações para células solares de perovskita