


MicroRNA-18b-3p exossômico derivado de células-tronco mesenquimais do cordão umbilical humano inibe a ocorrência de pré-eclâmpsia por direcionamento de LEP
Resumo
Exossomos derivados de células-tronco mesenquimais de cordão umbilical humano (hucMSCs) expressando microRNAs têm sido destacados em doenças humanas. No entanto, o mecanismo molecular detalhado de miR-18b-3p exossômico derivado de hucMSCs na pré-eclâmpsia (PE) permanece uma investigação mais aprofundada. Nosso objetivo foi investigar o efeito de exossomos e miR-18b-3p / leptina (LEP) na ocorrência de PE. A morfologia do hucMSC e hucMSC-exossomos (Exos) foi identificada. Os exossomos foram infectados com diferentes lentivírus expressando miR-18b-3p para explorar o papel do miR-18b-3p em PE. O modelo de rato PE foi estabelecido por injeção intraperitoneal de N éster metílico de -nitro-l-arginina. A expressão de LEP e miR-18b-3p foi testada em tecidos de placenta de rato PE. Além disso, o efeito dos exossomos na expressão de LEP e miR-18b-3p foi detectado. Foram detectados a pressão arterial sistólica (PAS), proteinúria, fatores inflamatórios, peso do feto de rato e placenta e apoptose celular em ratos com PE. Finalmente, a relação entre miR-18b-3p e LEP foi verificada usando ensaio de gene repórter de luciferase dupla e ensaio pull-down de RNA. Os exossomos, restaurando miR-18b-3p ou inibindo LEP reduziram a SBP e a proteinúria de ratos PE, bem como aumentaram o peso de ratos fetais e placenta, diminuíram os níveis séricos de fatores inflamatórios, bem como suprimiram células apoptóticas de ratos PE, exercendo um efeito supressor na progressão de PE. miR-18b-3p foi diminuído e LEP foi aumentado nos tecidos da placenta de ratos PE. LEP era o gene alvo direto de miR-18b-3p. A regulação positiva de miR-18b-3p ou o tratamento dos exossomos suprimiu a expressão de LEP e reduziu a ocorrência de PE, enquanto a regulação negativa de miR-18b-3p teve efeitos contrários. A LEP regulada para baixo reverteu o efeito da redução de miR-18b-3p em ratos com PE. O miR-18b-3p exossômico derivado de HucMSCs direciona a LEP para participar na ocorrência e no desenvolvimento de PE. Este estudo pode fornecer uma nova base teórica para o mecanismo e investigação do PE.
Introdução
A pré-eclâmpsia (PE), caracterizada por proteinúria e hipertensão [1], é uma das principais causas de mortalidade e morbidade fetal e materna na gravidez humana [2]. A etiologia e a patogênese da EP não são claras [3], que foi relatada como associada à invasão trofoblástica anormal resultando em disfunção endotelial materna, má perfusão placentária crônica e hipertensão com resultados adversos [4]. Com exceção do parto fetal e placentário, não há terapia específica para EP [5]. Portanto, é urgente explorar os alvos terapêuticos para melhorar o prognóstico desta doença.
O cordão umbilical humano (huc) é uma fonte adequada de células-tronco mesenquimais (MSCs) que secretam uma variedade de fatores tróficos e citocinas, bem como apresentam fortes capacidades antiinflamatórias e imunomoduladoras [6]. Um estudo verificou que o PE acelera a expressão do marcador neuroglial em MSCs derivadas de geléia de Wharton do cordão umbilical [7]. Um efeito protetor de exossomos de hucMSCs (Exos) na morfologia placentária e angiogênese em ratos PE também foi relatado [8]. Os exossomos são pequenas vesículas secretoras (50-100 nm) que medeiam a comunicação entre as células no microambiente tumoral por meio do encapsulamento e transmissão de fatores carcinogênicos para locais distantes ou para células circundantes pela circulação [9]. Um estudo revelou o dano das funções vasculares e complicações induzidas pela transferência eficaz de sFlt-1 e sEng para as células endoteliais em pacientes com PE por exossomos [10]. MicroRNAs (miRNAs) são RNAs endógenos não codificantes com um comprimento de 18–25 nucleotídeos e regulam a expressão do gene no nível pós-transcricional [11]. Os dados em um estudo relataram que a expressão do miR-18b afeta a invasão celular, a viabilidade e a migração das células trofoblásticas em PE [12]. Além disso, Wu et al. propuseram que o miR-18b atenua a proliferação em células endoteliais retinais humanas induzidas por alto teor de glicose, o que pode oferecer uma nova visão sobre a compreensão do mecanismo da patogênese da retinopatia diabética [13]. No entanto, o papel do miR-18b-3p exossômico derivado de hucMSC em PE permanece desconhecido. A leptina (LEP) tem efeitos pleiotrópicos na diferenciação / proliferação celular e imunidade dos estados fisiológicos e emergiu principalmente dos adipócitos, além de outros tecidos, incluindo a placenta [14]. Um estudo verificou que a metilação anormal do promotor LEP está envolvida na progressão do PE [15]. Outro estudo sugeriu que a placenta é o principal local de expressão da LEP na gravidez [16]. No entanto, a relação de ligação entre miR-18b-3p e LEP ainda é indescritível. Portanto, nosso objetivo foi explorar o papel de miR-18b-3p exossômico derivado de hucMSC em PE com o envolvimento de LEP, e inferimos que miR-18b-3p exossômico derivado de hucMSC pode inibir a progressão de PE por meio de direcionamento de LEP.
Materiais e métodos
Aprovação ética
O estudo foi aprovado pelo Comitê de Revisão Institucional do Hospital Popular da Universidade de Wuhan. Todos os participantes assinaram um termo de consentimento livre e esclarecido. Todos os experimentos com animais estavam de acordo com o Guia para o Cuidado e Uso de Animais de Laboratório dos Comitês Internacionais do Hospital Popular da Universidade de Wuhan.
Isolado, cultura e identificação de HucMSCs
O cordão umbilical fetal entregue por puérpera saudável foi coletado e cortado em picado e filtrado com peneira, em seguida, misturado com solução salina tamponada com fosfato (PBS). Os tecidos do cordão umbilical foram centrifugados a 1500 r / min por 5 min com 10 cm de raio centrífugo. Os tecidos foram suspensos com meio Eagle modificado por Dulbecco (DMEM) / F12 contendo 10% de soro fetal bovino (FBS) e transferidos para um frasco de cultura. Os líquidos foram trocados após 4 dias e, a seguir, trocados uma vez a cada 3 dias. As células foram sub-cultivadas quando a confluência atingiu cerca de 90%. O crescimento aderente e a morfologia de hucMSCs foram observados ao microscópio óptico. As células foram coradas com solução de coloração com óleo vermelho O (Beyotime Institute of Biotechnology, Shanghai, China) para detectar a diferenciação osteogênica de hucMSCs e tingidas com solução de coloração de fosfatase alcalina (ALP) (Beyotime) para detectar a diferenciação adipogênica de hucMSCs. Um citômetro de fluxo (Beckman Coulter Life Sciences, Brea, CA, EUA) foi adotado para testar CD73, CD166 (ambos 1:10, BD Biosciences, Franklin Lakes, NJ, EUA) e CD105 (1:20, AbD Serotec, Oxford, REINO UNIDO).
Extração e identificação de HucMSC-Exos
Os hucMSCs bem crescentes foram cultivados. O sobrenadante foi coletado e centrifugado a 28.500 r / min por 1 h com raio de centrífuga de 10 cm. O sobrenadante foi descartado, e as células fixadas com glutaraldeído 2% e ácido ósmico 1%, desidratado com etanol, imerso em óxido de propileno, seco por 2 h, embebido em Epon812 e fatiado. As fatias foram coradas com urânio e chumbo, respectivamente. Finalmente, os exossomos foram observados ao microscópio eletrônico. O detector Nanosight (Malvern Instruments, Malvern, UK) foi utilizado para detectar imagens de movimento browniano de nanopartículas de exossomos e seu tamanho. Os marcadores de superfície de hucMSC-Exos foram identificados por Western blot assay, e os resultados mostraram que hucMSC-Exos expressa CD9, CD81 e CD63.
Método de infecção por lentivírus
HucMSC foi infectado com lentivírus contendo baixa expressão do vetor miR-18b-3p e baixa expressão do controle negativo do vetor miR-18b-3p (NC) (Shanghai GenePharma Co, Ltd, Shanghai, China). Finalmente, o hucMSC-antagomir NC e o hucMSC-miR-18b-3p antagomir expresso de forma estável foram obtidos. As células foram cultivadas por 48 h, e o sobrenadante foi coletado e centrifugado com ultracentrifugação para obter o Exos-antagomir NC e o antagomir Exos-miR-18b-3p correspondentes.
Animais Experimentais
Ratos Wistar (pesando 200-250 g, idade de 8 w, independentemente do sexo) em um nível saudável e com maturidade sexual foram selecionados (o Experimental Animal Center da Wuhan University, Wuhan, China). Os ratos foram alimentados em um sistema de barreira com temperatura de 18-28 ° C, umidade relativa de 40-70% e dieta e água adequadas.
Estabelecimento de modelos de PE de ratos
O modelo de PE de rato foi estabelecido por injeção intraperitoneal de 50 mg / kg de inibidor de óxido nítrico sintetase, éster metílico de N (G) -nitro-l-arginina (L-NAME, Beyotime) com referência a um artigo [17]. O estabelecimento bem-sucedido do modelo de PE foi baseado no aumento da pressão arterial com 20 mmgHg e acima de 115 mmHg, bem como no aumento da proteinúria.
Agrupamento de animais
O rato fêmea e o rato macho coabitaram aleatoriamente em 1:1, e os dois ratos foram mantidos em uma gaiola especial individual às 17h - 18h. o dia anterior. O esperma nas secreções vaginais das ratas foi observado por tampão vaginal e microscópio no dia seguinte. Se o resultado fosse positivo na mesma hora, o dia era registrado como 0º dia de gestação. Do 13º rd dia de gestação, os ratos foram divididos em 6 grupos (10 ratos em cada grupo):grupo normal (a mesma quantidade de solução salina normal foi injetada por via intraperitoneal do dia 13 ao dia 20 de gestação), grupo PE (L-NAME [50 mg / kg por dia] foi injetado intraperitonealmente do dia 13 ao dia 20 de gestação, e 20 μL de solução salina normal foram injetados na placenta no dia 16 ao dia 19 de gestação), grupo PE + miR-NC (L-NAME [50 mg / kg por dia] foi injetado intraperitonealmente do dia 13 ao dia 20 de gestação, e 20 μL de 4 nmol miR-NC foram injetados na placenta no dia 16 ao dia 19 de gestação), grupo PE + miR-18b-3p agomir (L-NAME [50 mg / kg por dia] foi injetado intraperitonealmente do dia 13 ao dia 20 de gestação, e 20 μL de 4 nmol de miR-18b-3p agomir foram injetados na placenta do dia 16 ao dia 19 de gestação) , PE + miR-18b-3p antagomir grupo (L-NAME [50 mg / kg por dia] foi injetado intraperitonealmente do dia 13 ao dia 20 de gestação, e 20 μL de 4 nmol miR-18b-3p antagomir foram injetados no p lacenta no dia 16 ao dia 19 de gestação), PE + miR-18b-3p antagomir + pequeno RNA interferente (si) -LEP grupo (L-NAME [50 mg / kg por dia] foi injetado por via intraperitoneal do dia 13 ao dia 20 de gestação, e 20 μL de 4 nmol de antagomir miR-18b-3p e si-LEP foram injetados na placenta no dia 16 ao dia 19 de gestação) e grupo PE + si-LEP (L-NAME [50 mg / kg por dia] foi injetado intraperitonealmente do dia 13 ao dia 20 de gestação, e 20 μL de 4 nmol si-LEP foram injetados na placenta no dia 16 ao dia 19 de gestação). Os ratos foram tratados com exossomos e exossomos portadores de lentivírus. Os ratos foram divididos em 5 grupos (10 ratos em cada grupo):grupo normal (a mesma quantidade de solução salina normal foi injetada intraperitonealmente do dia 13 ao dia 20 de gestação), grupo PE (L-NAME (50 mg / kg por dia ) foi injetado intraperitonealmente do dia 13 ao dia 20 de gestação, e 20 μL de solução salina normal foram injetados na placenta no dia 16 ao dia 19 de gestação), grupo PE + Exos (L-NAME (50 mg / kg por dia) foi injetado intraperitonealmente do dia 13 ao dia 20 de gestação, e 20 μL de Exos (80 μg de exossomos foram suspensos em 20 μL de solução salina normal) foram injetados na placenta no dia 16 ao dia 19 de gestação), PE + Exos-antagomir NC grupo (L-NAME (50 mg / kg por dia) foi injetado intraperitonealmente do dia 13 ao dia 20 de gestação, e 20 μL de Exos-antagomir NC (80 μg de exossomos foram suspensos em 20 μL de solução salina normal) foram injetados na placenta no dia 16 ao dia 19 de gestação) e grupo PE + Exos-miR-18b-3p antagomir (L-NAME (50 mg / kg por dia) foi injetado por via intraperitoneal a partir do dia 13 ao dia 20 de gestação e 20 μL de Exos-miR-18b-3p antagomir (80 μg de exossomos foram suspensos em 20 μL de solução salina normal) foram injetados na placenta no dia 16 ao dia 19 de gestação).
Detecção de pressão arterial sistólica (PAS) e determinação de proteinúria 24 h
A pressão dos ratos foi medida pela medição da pressão sanguínea da artéria cauda dos ratos. A PAS do punho da cauda de todas as ratas grávidas foi medida no 10º, 13º, 16º e 19º dia de gestação usando o detector de pressão da artéria cauda de rato (Tensys (R) Medical Inc., San Diego, CA, EUA). A pressão foi medida 3 vezes em um curto espaço de tempo; a seguir, o valor médio foi tomado como a pressão arterial.
No caso da dieta livre e água, a urina de 24 horas das ratas grávidas foi coletada nos dias 10, 13, 16 e 19 de gestação, e o teor de proteína foi detectado no departamento de nefrologia do Hospital Popular da Universidade de Wuhan.
Amostra de coleta
As ratas grávidas foram anestesiadas com pentobarbital sódico a 3% no 21º dia de gestação. O sangue periférico dos ratos foi preservado, centrifugado para a coleta do soro e armazenado em geladeira a -20 ° C para standby. Em seguida, o feto do rato e a placenta foram retirados por cesariana, a membrana fetal e o cordão umbilical conectado foram removidos e o cordão umbilical conectado ao feto do rato foi cortado. A placenta e o feto do rato foram colocados em gaze asséptica para secar sangue e líquido amniótico, respectivamente, e a seguir colocados na balança analítica para pesar o peso. Uma parte dos tecidos placentários foi fixada com paraformaldeído 4%, desidratado com etanol, depurado com xileno, embebido com parafina e continuamente fatiado (5 μm) para coloração de hematoxilina-eosina (HE) e desoxinucleotidil transferase mediada por desoxinucleotidil transferase deoxiuridina trifosfato-biotina coloração TUNEL (nick end-labeling). O restante foi armazenado a -80 ° C para detecção da reação em cadeia da polimerase quantitativa da transcrição reversa (RT-qPCR), análise de Western blot e ensaio imunoenzimático (ELISA).
ELISA
Os teores de fator de necrose tumoral-α (TNF-α), interleucina (IL) -1β e IL-6 no soro foram testados por ELISA. As concentrações de TNF-α, IL-1β e IL-6 foram determinadas seguindo as instruções do kit (R&D Systems, Minneapolis, MN, EUA). Os valores de densidade óptica (DO) (490 nm) foram testados por um leitor de microplaca (Thermo Fisher Scientific, MA, EUA). A curva padrão correspondente foi obtida usando o valor de DO como abcissa e a concentração da amostra padrão correspondente como ordenada. As concentrações de TNF-α, IL-1β e IL-6 foram calculadas a partir da curva padrão.
Coloração HE
As amostras de parafina dos tecidos da placenta foram depuradas em xilol, desidratadas por gradiente de álcool convencional, coradas com hematoxilina, diferenciadas por álcool ácido clorídrico 1% e devolvidas ao azul por água amoniacal 1%. Em seguida, os tecidos foram contracorados com solução de eosina 1%, desidratados (75%, 90%, etanol 95%, respectivamente, álcool etílico absoluto) e clarificados por xileno, secos, bloqueados e observados ao microscópio eletrônico.
Coloração TUNEL
As seções embebidas em parafina foram rotineiramente desparafinadas e desidratadas de acordo com as instruções e, em seguida, a apoptose foi detectada pelo TUNEL Kit (Nanjing Kejin Biotechnology Co., Ltd., Jiangsu, China). 4,6-diamino-2-fenilindol (Shanghai Baitai Biotechnology Co., Ltd., Shanghai, China) foi usado para observar células TUNEL-positivas usando um microscópio de fluorescência (Nikon, Tóquio, Japão) [18].
RT-qPCR
Os tecidos da placenta foram pesados. Por 50-100 mg, os tecidos da placenta foram adicionados com 1 mL de TRIzol (Invitrogen, Carlsbad, Califórnia, EUA) e completamente dissolvidos. Os tecidos foram anexados com 200 μL de clorofórmio e centrifugados a 4 ° C, 12.000 rpm para extrair o RNA total. A concentração e a pureza do RNA foram determinadas por espectrofotômetro de ácido nucléico de proteína DU-800 (Beckman). U6 e β-actina foram utilizados como controles de carga. Os primers de PCR foram projetados e compostos pela Shanghai Sangon Biotechnology Co. Ltd. (Shanghai, China). As sequências do iniciador estão listadas na Tabela 1. O RNA foi revertido para cDNA com base nas instruções do kit de transcrição reversa de RNA (Sangon). O PCR foi amplificado e os produtos foram verificados por eletroforese em gel de agarose. Os dados foram calculados por 2 −ΔΔCt método.
Ensaio de Western Blot
A proteína total dos tecidos da placenta foi extraída por ensaio de radioimunoprecipitação com tampão de lise celular (Beyotime). HucMSC-Exo foi utilizado para abstrair o tampão, que foi centrifugado a 14.000 rpm. O sobrenadante foi preservado para testar a expressão proteica da proteína marcadora exossômica (CD81, CD63 e CD9) no soro. A concentração de proteína foi determinada pelo kit de ácido bicinconínico (Beyotime, P0010). A amostra foi carregada de acordo com os resultados quantitativos da proteína, tratada com eletroforese em gel de dodecilsulfato de sódio-poliacrilamida a 10% e transferida para membrana. A membrana foi bloqueada com 5% de leite desnatado, sondado com anticorpos primários LEP, CD63, CD81, CD9 e β-actina (4 mL, 1:1000, Santa Cruz Biotechnology, Inc, Santa Cruz, CA, EUA), re-sondado com 4 mL de anticorpo secundário de cabra anti-IgG de coelho / peroxidase de rábano, exposto e desenvolvido. A β-Actina foi utilizada como referência interna. O valor de cinza foi analisado pelo software de análise gráfica de gel Image Lab.
Ensaio do gene Dual-Luciferase Reporter
O software de previsão on-line https://cm.jefferson.edu/ foi adotado para prever a relação alvo entre miR-18b-3p e LEP, bem como o sítio de ligação de miR-18b-3p e região não traduzida LEP 3′ (UTR ) A sequência da região promotora LEP 3′UTR contendo o sítio de ligação miR-18b-3p foi composta. O plasmídeo LEP 3′UTR de tipo selvagem (WT) e o tipo mutante LEP 3′UTR (MUT) foram construídos. Os plasmídeos recombinantes foram nomeados como LEP 3′UTR-WT e LEP 3′UTR-MUT, respectivamente. As células 293T cultivadas foram co-transfectadas com miR-18b-3p mimetizador e LEP 3′UTR-WT, miR-18b-3p mimetizador e LEP 3′UTR-MUT, mimetizador NC e LEP 3′UTR-WT, mimetizador NC e LEP 3′UTR-MUT por 30 h. Em seguida, as células 293T foram coletadas. A atividade da luciferase do vagalume e da renila em células foi medida por medições de luminescência de acordo com o kit de detecção de gene repórter de luciferase dupla (Promega, Madison, WI, EUA).
Ensaio pull-down de RNA
Sondas de RNA biotinilado (Bio-miR-NC, Bio-miR-18b-3p e Bio-miR-18b-3p-Mut) foram incubadas com o lisado de células 293T e extraídas usando grânulos magnéticos conjugados com estreptomicina antibiótico. O experimento foi realizado com base nas instruções dos kits pull-down de RNA magnético Pierce (Pierce, IL, EUA). O RNA foi eluído e purificado usando TRIzol (Pierce). O enriquecimento de LEP em complexo de RNA foi quantificado usando RT-qPCR como descrito anteriormente [19].
Análise estatística
Todos os dados foram explicados pelo software SPSS 21.0 (IBM Corp. Armonk, NY, EUA). Os dados de medição foram indicados como média ± desvio padrão. Os dados foram conduzidos por amostra independente t teste para comparações de dois grupos, enquanto as comparações entre vários grupos foram avaliadas por análise de variância unilateral (ANOVA) seguida pelo teste post hoc de Tukey. O critério de significância estatística foi estabelecido em p <0,05.
Resultados
Morfologia e identificação do HucMSC e HucMSC-Exos
As massas de tecido do cordão umbilical foram observadas ao microscópio invertido. Pode-se ver que as células rastejaram para fora da massa de tecido no dia 3; as células mostraram forma de fuso e filamentosos, bem como cresceram como uma colônia em cerca de 5 dias. Quando cultivadas até a passagem 3, a morfologia das células era fusiforme longa e uniforme e semelhante à morfologia dos fibroblastos, e o arranjo era regular (Fig. 1a). Após 2 w de diferenciação adipogênica de hucMSC, gotículas de lipídios foram formadas no citoplasma, e as gotículas de lipídios mostraram estrutura de Kranz sob o microscópio invertido (Fig. 1b), sugerindo que o hucMSC cultivado isolado tinha a capacidade de diferenciação adipogênica. Após 2 w de diferenciação osteogênica, um grande número de nódulos de cálcio castanhos pode ser visto sob um microscópio invertido (Fig. 1c), indicando que o hucMSC cultivado isolado tinha a capacidade de diferenciação osteogênica. Um citômetro de fluxo foi adotado para testar o imunofenótipo das células, e os resultados incluíram que as células superexpressaram os marcadores de superfície CD73, CD105 e CD166 de MSCs (Fig. 1d).
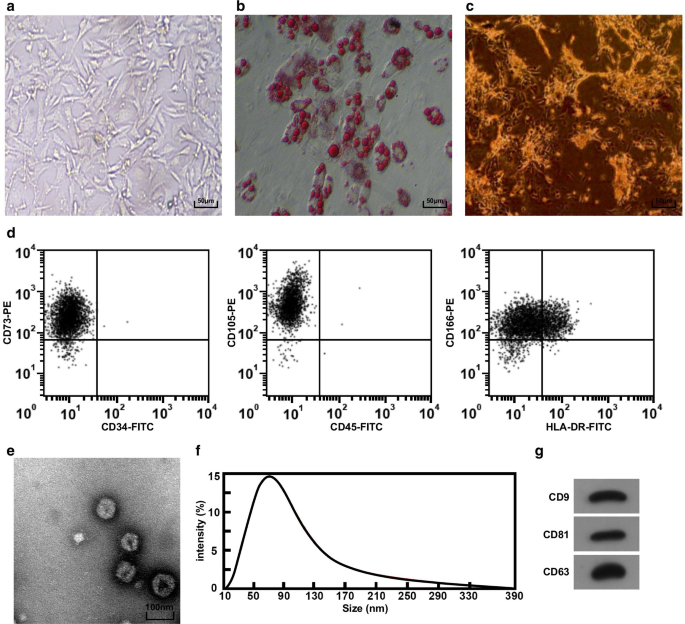
Morfologia e identificação do hucMSC e hucMSC-Exos. a A morfologia do hucMSC sob o microscópio invertido, b O hucMSC foi testado por coloração com óleo vermelho O. c hucMSC foi testado por coloração ALP. d A citometria de fluxo foi usada para detectar o imunofenótipo. e A forma e o tamanho de hucMSC-Exos observados por meio de um TEM. f Detecção da distribuição de tamanho de partícula de exossomos usando análise Nanosight. g A expressão da proteína de CD9, CD81 e CD63 em hucMSC-Exos foi detectada por ensaio de Western blot
A morfologia do hucMSC-Exos foi observada pelo TEM, e os resultados mostraram que os exossomos eram redondos ou ovais com baixa densidade central e coloração espessa em ambos os lados (Fig. 1e). A análise de nanosight foi utilizada para analisar o tamanho das partículas dos exossomos, e os resultados mostraram que o tamanho das partículas estava principalmente distribuído entre 40 e 100 nm, mais concentrado em torno de 80 nm (Fig. 1f). O ensaio de Western blot revelou que todos os marcadores de superfície CD81, CD63 e CD9 foram expressos em hucMSC-Exos (Fig. 1g).
Restaurar miR-18b-3p alivia as características patológicas de ratos com PE
Os resultados da PAS e da proteinúria de 24 horas apresentaram que:não houve diferença significativa na PAS e na proteinúria de 24 horas em 6 grupos antes da administração (10º dia de gestação). A PAS e a proteinúria de 24 horas no dia 19 de gestação não mostraram nenhuma diferença óbvia em ratos normais. Em ratos PE ou ratos PE tratados com miR-NC, miR-18b-3p agomir, miR-18b-3p antagomir, miR-18b-3p antagomir + si-LEP, si-NC ou si-LEP, SBP e 24-h a proteinúria começou a aumentar no dia 13 de gestação. Não houve diferença distinta de PAS e proteinúria de 24 horas no dia 16 e no dia 19 de gestação em ratas PE tratadas com miR-18b-3p agomir e si-LEP. Ratas PE tiveram aumento da PAS e proteinúria de 24 horas no dia 19 de gestação; este aumento foi reduzido pela elevação do miR-18b-3p, mas ainda mais potenciado pela inibição do miR-18b-3p; A redução da LEP anulou o papel da regulação negativa do miR-18b-3p na PAS e na proteinúria de 24 horas no dia 19 de gestação em ratos com PE (Fig. 2a, b).
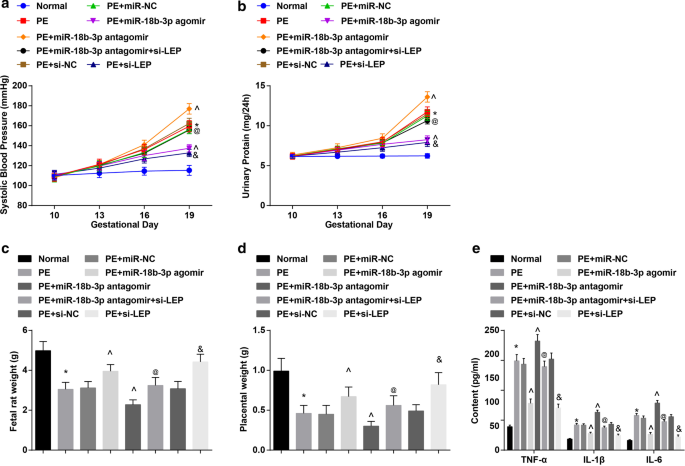
Restaurar miR-18b-3p alivia as características patológicas de ratos PE. a Resultados da SBP em ratos. b Resultados da proteinúria de 24 horas em ratos. c Alterações de peso em fetos de ratos. d Alterações do peso da placenta em ratos. e Alterações de fatores inflamatórios no soro foram detectados por ELISA. n =10, * p <0,05 versus o grupo normal. ^ p <0,05 versus o grupo PE + miR-NC. @ p <0,05 versus o grupo PE + miR-18b-3p antagomir. e p <0,05 versus o grupo PE + si-NC. Os dados de medição foram representados como média ± desvio padrão, e as comparações entre vários grupos foram avaliadas por ANOVA de uma via seguida pelo teste de Tukey
O peso do feto do rato e da placenta foi reduzido nos ratos com PE; miR-18b-3p regulado positivamente ou LEP regulado negativamente aumentou, enquanto miR-18b-3p regulado negativamente diminuiu o peso do feto de rato e da placenta em ratos PE; O silenciamento da LEP reverteu o efeito da inibição do miR-18b-3p no peso do feto do rato e da placenta em ratos PE (Fig. 2c, d).
Fatores inflamatórios no soro de ratos PE foram detectados. Verificou-se que os teores de TNF-α, IL-1β e IL-6 aumentaram nos ratos PE; elevação de miR-18b-3p ou inibição de LEP suprimida, enquanto a redução de miR-18b-3p promoveu os conteúdos de TNF-α, IL-1β e IL-6; o efeito do miR-18b-3p inibido nos conteúdos de TNF-α, IL-1β e IL-6 foi anulado pela depleção de LEP (Fig. 2e).
O miR-18b-3p superexpresso melhora a alteração histopatológica dos tecidos da placenta de ratos com PE
Em ratos normais, a vilosidade placentária era rica em vasos sanguíneos e tinha uma estrutura clara, os sincitiotrofoblastos eram os principais trofoblastos nas vilosidades placentárias e havia menos citotrofoblastos. Em ratos PE ou ratos PE tratados com miR-NC, miR-18b-3p antagomir, si-NC ou miR-18b-3p antagomir + si-LEP, o número de vilosidades placentárias diminuiu, a estrutura estava turva e atrofiada, algumas vilosidades foram realizadas necrose fibrinóide, e o número de nódulos de sincitiotrofoblastos nas vilosidades placentárias aumentou, sendo a maioria das vilosidades imaturas. O número de trofócitos foi reduzido e as alterações patológicas foram aliviadas em ratos PE tratados com miR-18b-3p agomir e si-LEP (Fig. 3a).
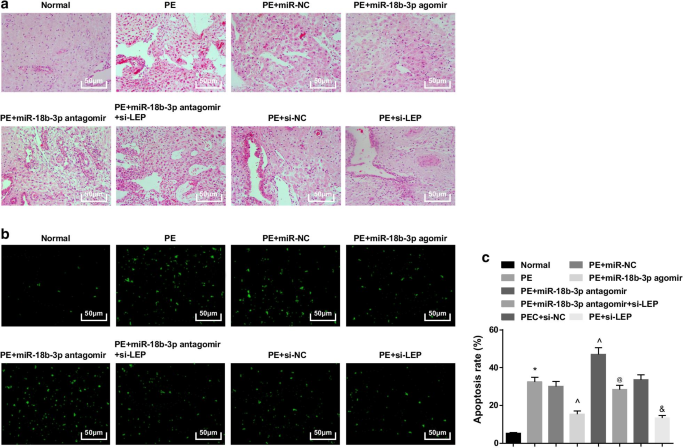
O miR-18b-3p superexpresso melhora a mudança patológica e suprime as células apoptóticas dos tecidos da placenta em ratos PE. a A coloração HE foi utilizada para testar características patológicas dos tecidos da placenta. b A coloração TUNEL foi implementada para determinar células apoptóticas de tecidos da placenta em ratos PE. c A taxa de apoptose celular foi detectada por coloração TUNEL. n =10, * p <0,05 versus o grupo normal. ^ p <0,05 versus o grupo miR-NC. @ p <0,05 versus o grupo antagomir miR-18b-3p. e p <0,05 versus o grupo si-NC. Os dados de medição foram representados como média ± desvio padrão, e as comparações entre vários grupos foram avaliadas por ANOVA de uma via seguida pelo teste de Tukey
A coloração de TUNEL sugeriu que um pequeno número de células apoptóticas pode ser visto. Os ratos PE apresentaram aumento das células apoptóticas, que foram reduzidas pela elevação do miR-18b-3p e silenciamento da LEP, e foram posteriormente aumentadas pela inibição do miR-18b-3p; O silenciamento de LEP também reverteu o efeito da inibição de miR-18b-3p no número de células apoptóticas em ratos PE (Fig. 3b, c).
Tomados em conjunto, os ratos com miR-18b-3p regulado positivamente ou LEP inibido tiveram um grau diminuído de progressão de PE na histologia, e LEP silenciada poderia abolir o efeito terapêutico do miR-18b-3p inibido.
miR-18b-3p é regulado para baixo, enquanto LEP é regulado para cima em tecidos de placenta de rato PE e miR-18b-3p alvos LEP
Com base nos resultados acima, a regulação negativa de LEP reverteu o efeito terapêutico da regulação negativa de miR-18b-3p em ratos PE em patologia e histologia; assim, formulamos a hipótese de que miR-18b-3p pode estar relacionado a LEP.
O ensaio de Western blot e RT-qPCR revelaram que ratos PE diminuíram miR-18b-3p e aumentaram os níveis de expressão de LEP; o tratamento de miR-18b-3p agomir regulou positivamente miR-18b-3p e diminuiu a regulação de LEP em ratos PE, enquanto o tratamento de miR-18b-3p antagomir aumentou a expressão de LEP; O silenciamento de LEP reverteu o efeito promotor da redução de miR-18b-3p na expressão de LEP em ratos PE (Fig. 4a-c).
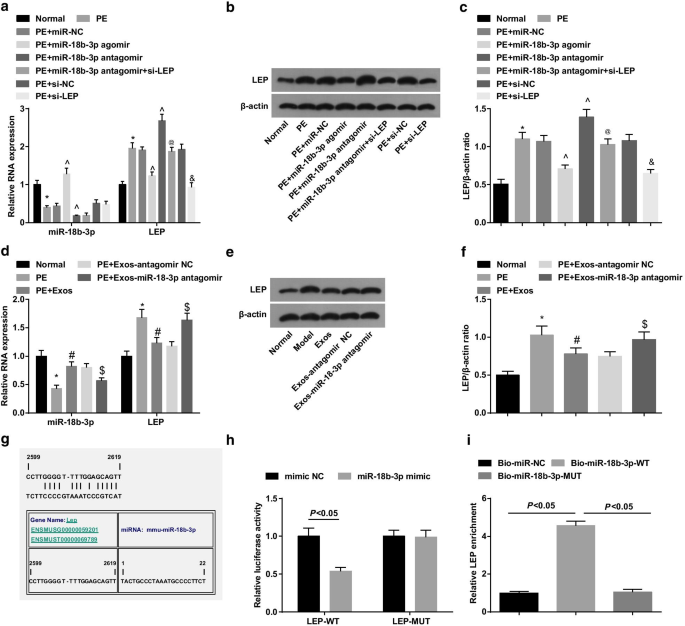
miR-18b-3p é regulado negativamente e LEP é regulado positivamente em tecidos da placenta de ratos PE. a A expressão de miR-18b-3p e mRNA de LEP em tecidos da placenta foi detectada usando RT-qPCR. b Faixa de proteína da LEP em tecidos da placenta. c O ensaio de Western blot foi conduzido para detectar a expressão da proteína LEP em tecidos da placenta. d A expressão de miR-18b-3p e mRNA de LEP em tecidos da placenta após o tratamento com exossomo foi detectada usando RT-qPCR. e Faixa de proteína da LEP em tecidos da placenta após tratamento com exossomo. f O ensaio de Western blot foi realizado para detectar a expressão da proteína LEP após o tratamento com exossomo. g Os sítios de ligação de miR-18b-3p e LEP previstos por um software online. h A relação alvo entre miR-18b-3p e LEP verificada por ensaio de gene repórter de luciferase dupla. eu relação de direcionamento entre miR-18b-3p e LEP verificada por ensaio pull-down de RNA. n =10, * p <0,05 versus o grupo normal. ^ p <0,05 versus o grupo miR-NC. @ p <0,05 versus o grupo antagomir miR-18b-3p. e p <0,05 versus o grupo si-NC. # p <0,05 versus o grupo PE. $ p <0,05 versus o grupo PE + Exos-antagomir NC. Os dados de medição foram representados como média ± desvio padrão, e as comparações entre vários grupos foram avaliadas por ANOVA de uma via seguida pelo teste de Tukey
O ensaio de Western blot e RT-qPCR foram usados para explorar o papel dos exossomos em ratos PE. Os resultados mostraram que os exossomos regularam positivamente o miR-18b-3p e negaram a LEP em ratos com PE, indicando o efeito supressor dos exossomos no desenvolvimento de PE. Além disso, os exossomos que transportam miR-18b-3p antagomir induziram a regulação negativa de miR-18b-3p e a regulação positiva de LEP em ratos PE (Fig. 4d-f).
A relação alvo entre miR-18b-3p e LEP foi prevista pelo software de previsão online de bioinformática https://cm.jefferson.edu/ (Fig. 4g). O ensaio do gene repórter de luciferase dupla sugeriu que o mimetizador de miR-18b-3p diminuiu a atividade da luciferase de LEP 3′UTR-WT, embora não impusesse nenhum impacto na de LEP 3′UTR-MUT (Fig. 4h). Além disso, o ensaio de pull-down de RNA revelou que o enriquecimento de LEP foi aumentado em miR-18b-3p biotinilado por WT (Fig. 4i). Essas descobertas indicaram que LEP é um gene alvo de miR-18b-3p.
hucMSC-Exos Atenua as características patológicas de ratos com PE
Os resultados da PAS e 24 h mostraram que não houve diferença significativa na PAS e na proteinúria 24 h em 5 grupos antes da administração (10º dia de gestação). A PAS e a proteinúria de 24 horas no dia 19 de gestação não mostraram diferenças distintas em ratas normais. Em ratas PE, a PAS e a proteinúria de 24 horas começaram a aumentar no 13º dia de gestação. Não houve diferença distinta de PAS e proteinúria de 24 horas no dia 16 e no dia 19 de gestação em ratas PE tratadas com hucMSC-Exos e hucMSC-Exos transmitindo antagomir NC. A PAS e a proteinúria de 24 horas aumentaram no dia 19 de gestação nas ratas PE, enquanto o aumento foi reduzido pela injeção de hucMSC-Exos. A inibição de miR-18b-3p reverteu o efeito de hucMSC-Exos na PAS e na proteinúria de 24 h no dia 19 de gestação em ratos PE (Fig. 5a, b).
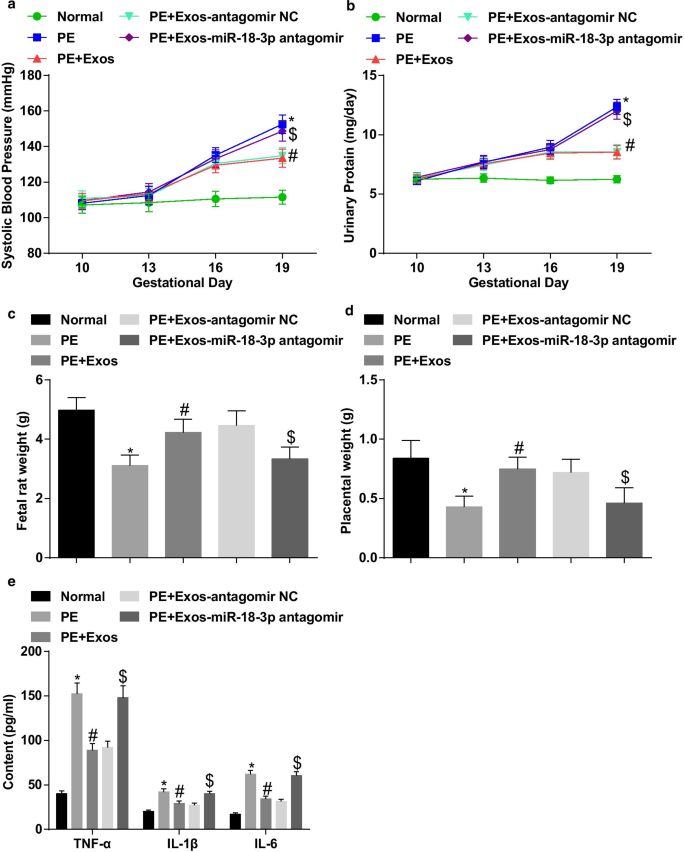
hucMSC-Exos atenua características patológicas de ratos PE. a Resultados da SBP em ratos após tratamento com exossomo. b Resultados da proteinúria de 24 horas em ratos após tratamento com exossomo. c Alterações de peso em fetos de ratos após tratamento com exossomo. d Alterações do peso da placenta em ratos após tratamento com exossomo. e As alterações dos fatores de inflamação após o tratamento com exossomo no soro foram determinadas por ELISA. n =10, * p <0,05 versus o grupo normal. # p <0,05 versus o grupo PE. $ p <0,05 versus o grupo PE + Exos-antagomir NC. Os dados de medição foram representados como média ± desvio padrão, e as comparações entre vários grupos foram avaliadas por ANOVA de uma via seguida pelo teste de Tukey
O peso do feto do rato e da placenta foi medido, e descobrimos que os ratos PE tinham peso reduzido do feto do rato e da placenta; A regulação negativa de miR-18b-3p aboliu o papel de hucMSC-Exos no peso de fetos de rato e placenta em ratos PE (Fig. 5c, d).
Fatores inflamatórios no soro foram detectados por ELISA. Os teores de TNF-α, IL-1β e IL-6 aumentaram notavelmente em ratos com PE. O tratamento com exossomos diminuiu os conteúdos de TNF-α, IL-1β e IL-6 no soro de ratos PE, que foram aumentados pela injeção de exossomos que inibem miR-18b-3p (Fig. 5e).
Os exossomos alivia as alterações patológicas e inibem a apoptose dos tecidos da placenta de ratos com PE
Em ratos normais, as vilosidades placentárias eram abundantes nos vasos sanguíneos com uma estrutura clara, os sinciciotrofoblastos eram o trofoblasto principal nas vilosidades placentárias e havia menos citotrofoblastos. Nos ratos PE e ratos PE tratados com hucMSC-Exos-miR-18b-3p-antagomir, o número de vilosidades placentárias reduziu, a estrutura estava turva e atrofiada, algumas vilosidades apresentavam necrose fibrinóide e o número de nódulos de sinciciotrofoblastos na placenta as vilosidades aumentaram e a maioria das vilosidades estava imatura. A mudança patológica foi melhorada nos ratos PE tratados com hucMSC-Exos ou hucMSC-Exos-antagomir NC contra os ratos PE e ratos PE tratados com hucMSC-Exos-miR-18b-3p antagomir (Fig. 6a).
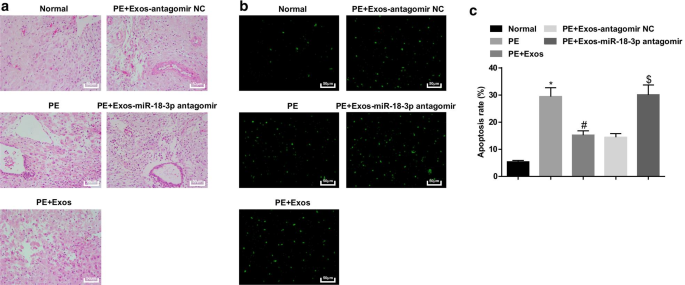
Os exossomos aliviam as alterações patológicas e diminuem as células apoptóticas dos tecidos da placenta em ratos PE. a A coloração HE foi utilizada para testar características patológicas dos tecidos da placenta em ratos PE após tratamento com exossomo. b A coloração TUNEL foi implementada para determinar células apoptóticas de tecidos da placenta em ratos PE após tratamento com exossomo. c A taxa de apoptose celular foi detectada por coloração TUNEL. n =10, * p <0,05 versus o grupo normal. # p <0,05 versus o grupo PE. $ p <0,05 versus o grupo PE + Exos-antagomir NC. Os dados de medição foram representados como média ± desvio padrão, e as comparações entre vários grupos foram avaliadas por ANOVA de uma via seguida pelo teste de Tukey
A coloração de TUNEL indicou que em ratos normais, um pequeno número de células apoptóticas pode ser visto. Ratos PE tinham células apoptóticas aumentadas e miR-18b-3p reduzido reverteu os impactos de hucMSC-Exos no número de células apoptóticas em tecidos de placenta de ratos PE (Fig. 6b, c).
Discussão
PE é um distúrbio multissistêmico da gravidez caracterizado por proteinúria e hipertensão ou outras condições adversas e está relacionado a uma ampla gama de disfunção endotelial materna [20]. Foi relatado que o hucMSC-Exo melhorou a morfologia do tecido placentário em ratos PE por meio da supressão da apoptose celular e facilitação da angiogênese no tecido placentário de uma maneira dependente da dose [8]. Um estudo relatou que a expressão de miR-18b afetou a migração celular, a viabilidade e a invasão em PE [12]. Além disso, foi verificado aumento da concentração materna de LEP e hipometilação da LEP na placenta no início da PE [21]. O estudo atual foi projetado para explorar o efeito dos exossomos e do LEP direcionado ao miR-18b-3p na ocorrência de PE. As descobertas neste estudo revelaram que miR-18b-3p exossômico derivado de hucMSC inibiu a progressão de PE ao reduzir LEP.
Com base em nossos achados, o miR-18b-3p foi reduzido e o LEP elevado nos tecidos da placenta de ratos com PE. Semelhante ao nosso estudo, a expressão de mRNA de miR-18b foi marcadamente suprimida em tecidos placentários com PE em relação aos tecidos placentários normais [12]. Além disso, um estudo revelou que o conteúdo de miR-18b foi drasticamente reduzido em tecidos de melanoma maligno em comparação com seus tecidos adjacentes não tumorais correspondentes [22]. Outro estudo verificou que a expressão de LEP placentária aumentou em PE pré-termo em comparação com controles [23]. Além disso, um estudo mostrou que a expressão de LEP estava obviamente aumentada em placentas pré-eclâmpticas [15]. Esta literatura forneceu uma base teórica para explorarmos a expressão anormal de miR-18b-3p e LEP em PE. Além disso, foi previsto usando um software de bioinformática que LEP foi direcionado por miR-18b-3p, e essa relação de direcionamento foi posteriormente confirmada com ensaio de gene repórter de luciferase dupla em nossa pesquisa. Um estudo relatou que LEP é um alvo para todos os três miRNAs (miR-1301, miR-223 e miR-224) em PE de início precoce [16]. Outro estudo demonstrou que LEP diminuiu a expressão de miR-93 na osteoartrite e na artrite reumatóide [24]. No entanto, a ligação entre miR-18b-3p e LEP em doenças humanas, principalmente em PE, permanece pouco estudada, o que é a novidade deste estudo. Além disso, um resultado emergente de nosso estudo relatou que os exossomos aumentaram o miR-18b-3p e diminuíram a LEP nos tecidos da placenta de PE. Foi documentado anteriormente que a expressão de miR-18b-5p foi notavelmente aumentada em exossomos plasmáticos de câncer colorretal [25], enquanto a relação entre hucMSC-Exos e miR-18b-3p / LEP em PE precisa de um estudo mais aprofundado.
Além disso, o achado de nossa investigação mostrou que o miR-18b-3p restaurado reduziu a PAS e a proteinúria de 24 h de ratos com PE, aumentou o peso da placenta, diminuiu os conteúdos de TNF-α, IL-1β e IL-6 no soro e nos tecidos da placenta bem como suprimida a apoptose celular. Esses dados indicaram que a elevação do miR-18b-3p contribui para o alívio dos sintomas e alterações patológicas na EP. Foi demonstrado que a suprarregulação estável de miR-18b produziu atividade inibidora de tumor eficaz, como inibir a viabilidade celular de melanoma, induzindo apoptose e reduzindo o crescimento tumoral in vivo [26]. Outro resultado em nosso estudo foi que o hucMSC-Exos reduziu a PAS e a proteinúria de 24 horas de ratos com PE, aumentou o peso da placenta, diminuiu o conteúdo de TNF-α, IL-1β e IL-6 no soro e nos tecidos da placenta, bem como células suprimidas apoptose. As descobertas do presente estudo revelaram que modelos de ratos PE tratados com exossomos apresentaram um aumento no número e na qualidade dos fetos, na qualidade da placenta, mas a apoptose celular foi significativamente reduzida [8]. Curiosamente, uma pesquisa anterior demonstrou que a adição de exossomos fetais bovinos diminuiu o conteúdo de macrófagos TNF-α e IL-6 [27]. Um estudo revelou que exossomos purificados suprimiram a produção de IL-1β em macrófagos estimulados por lipopolissacarídeo / nigericina [28]. Além disso, Nong et al. sugeriram que marcadores inflamatórios, como TNF-α e IL-6, diminuíram drasticamente após a administração de exossomos produzidos por células-tronco pluripotentes induzidas por humanos, MSCs [29]. Há um artigo descobrindo que a PAS foi acentuadamente elevada no grupo de mulheres que posteriormente desenvolveram PE [30, 31]. Foi demonstrado que os pacientes com PE foram positivamente associados com PAS e pressão arterial diastólica e proteinúria [32]. Além disso, um estudo recente forneceu uma prova de que a proteinúria aumentou com o avanço da gestação em mulheres com PE [33]. Um achado importante foi que as ratas do grupo de PE tinham TNF-α aumentado em relação ao grupo de prenhes normal [34]. Outro estudo verificou que o soro IL-6 e IL-1β estavam obviamente elevados em mulheres com PE em relação aos controles [35]. Os achados acima sugeriram que os pacientes com PE geralmente apresentavam PAS elevadas, proteinúria e níveis de fatores inflamatórios. Assim, pode-se inferir de nossos resultados que o miR-18b-3p exossômico derivado de hucMSC teve um efeito terapêutico no PE.
Conclusão
Em conclusão, nosso estudo fornece evidências de que exossomos derivados de hucMSCs regulam positivamente miR-18b-3p, que visa a LEP para suprimir o conteúdo de fatores inflamatórios e reduzir a taxa de apoptose celular em tecidos de placenta de rato com PE, inibindo assim a ocorrência de PE. Assim, miR-18b-3p exossômico pode ser um candidato potencial para o tratamento de PE via direcionamento de LEP. Esta pesquisa identificou o papel do miR-18b-3p exossômico derivado de hucMSC visando LEP durante o desenvolvimento de PE pela primeira vez, o que forneceu um novo insight para o tratamento de PE. No entanto, devido à limitação de pesquisas conhecidas, o estudo precisa ser monitorado rigorosamente e relatado de forma adequada em futuros ensaios clínicos.
Disponibilidade de dados e materiais
Não aplicável.
MicroRNA mediada por HDAC1-124-5p regula NPY para afetar habilidades de aprendizagem e memória em ratos com depressão
Investigações estruturais de ilhas com bicamadas de boro-carbono em escala atômica em cristal único de diamante altamente dopado com boro:origem da tensão de tração em etapas
Nanomateriais
- Como o cádmio prejudica o corpo humano?
- Neurônios artificiais podem ser tão eficientes quanto o cérebro humano
- A chegada da cadeia de suprimentos ‘autônoma’
- O Elemento Humano Crítico na Estratégia de Cibersegurança
- Na pandemia, o compromisso das fábricas com os direitos humanos está caindo
- O futuro da automação na indústria da aviação
- Sistema de segurança biométrica baseado no batimento cardíaco humano
- Delaware:a mistura perfeita entre humanos e IA
- O elemento humano da automação
- O fator humano na inspeção de metais