


A regulação negativa de microRNA-342-5p ou a regulação positiva de Wnt3a inibe a angiogênese e mantém a estabilidade da placa aterosclerótica em camundongos com aterosclerose
Resumo
Evidências demonstraram que o microRNA-342-5p (miR-342-5p) está implicado na aterosclerose (EA), mas pouco se sabe sobre seus mecanismos regulatórios intrínsecos. Aqui, nosso objetivo foi explorar o efeito de miR-342-5p direcionado a Wnt3a na formação de placas vulneráveis e angiogênese de AS. ApoE - / - camundongos foram alimentados com ração rica em gordura por 16 semanas para replicar o modelo de placa vulnerável AS. A expressão de miR-342-5p e Wnt3a em tecidos aórticos de AS foi detectada. A relação de destino entre miR-342-5p e Wnt3a foi verificada. Além disso, ApoE - / - camundongos foram injetados com miR-342-5p antagomir e vetor de superexpressão Wnt3a para testar suas funções nos níveis de lipídios séricos, citocinas relacionadas ao estresse inflamatório e oxidativo, estabilidade da placa aórtica e angiogênese em placa de camundongos AS. A expressão de miR-342-5p foi aumentada e a expressão de Wnt3a foi degradada em tecidos aórticos de camundongos AS e miR-342-5p diretamente direcionado a Wnt3a. A regulação positiva de Wnt3a ou a regulação negativa de miR-342-5p reduziu o conteúdo de lipídios no sangue, os níveis de estresse inflamatório e oxidativo, a vulnerabilidade da placa do tecido aórtico e inibiu a angiogênese na placa aórtica de camundongos AS. Estudos funcionais mostram que o esgotamento de miR-342-5p pode estabilizar a placa do tecido aórtico e reduzir a angiogênese na placa em camundongos AS por meio da restauração de Wnt3a.
Introdução
A aterosclerose (EA) é uma doença arterial relacionada à idade caracterizada pelo espessamento, estenose, endurecimento e formação de placas ateroscleróticas das artérias [1]. É a causa prevalente de morte e morbidade em países desenvolvidos [2]. Estudos histopatológicos de lesões ateroscleróticas humanas mostraram que o desenvolvimento e a ruptura da placa são caracterizados pela expansão do núcleo de lipídios / necrose, redução no número de células musculares lisas, infiltração de macrófagos e diminuição do conteúdo de colágeno [3]. Os principais elementos celulares da AS incluem hiperlipidemia, formação de células espumosas, diferenciação em macrófagos, recrutamento de monócitos e inflamação induzida [4]. Embora muitos medicamentos para o tratamento da EA tenham sido amplamente utilizados na clínica, alguns subgrupos de pacientes ainda apresentam alto risco de infarto do miocárdio, isquemia do miocárdio, insuficiência cardíaca e acidente vascular cerebral [5]. Portanto, uma exploração mais aprofundada dos potenciais mecanismos moleculares pode oferecer mais evidências para o tratamento de EA.
Um único microRNA (miRNA) pode regular simultaneamente vários alvos de genes [6]. O miR-342-5p é investigado para estar contido no agrupamento de miRNA 14q32 impresso, atua como uma molécula Notch a jusante inovadora [7] e modula várias vias angiogênicas, como a sinalização do fator de crescimento transformador β e o fator de crescimento endotelial vascular [8]. MicroRNAs imunomoduladores relacionados, como miR-342-5p, têm vários papéis importantes na regulação da progressão da aterosclerose [9]. Além disso, alguns miRNAs foram sugeridos para envolver na resolução de AS, como miR-155 e miR-217 [10, 11]. Um estudo relatou que miR-342-5p atua como um novo modulador para ativação de macrófagos em AS [12]. Outro estudo revelou que o miR-342-5p derivado de macrófagos facilita a EA e aumenta a estimulação inflamatória dos macrófagos [13]. Qu et al . descobriram que a expressão de Wnt3a é modulada negativamente por miR-342-5p em malformações anorretais [14], indicando que há uma relação de alvo entre miR-342-5p e Wnt3a. A sinalização Wnt desempenha um papel essencial durante a embriogênese para a modulação da polaridade celular, proliferação celular, formação de eixos e apoptose [15]. Wnt3a, um componente chave do gene mesoderma, desempenha um papel crucial no desenvolvimento embrionário [16]. Foi apresentado que a análise guiada por epigenoma do transcriptoma de macrófagos da placa durante a regressão AS revela a ativação da via de sinalização Wnt [17]. Além disso, um estudo relatou que Wnt3a modula a adesão e migração de células do músculo liso vascular que contribuem para a patogênese da EA e reestenose [18]. Portanto, este estudo explorou pela primeira vez o efeito de miR-342-5p direcionado Wnt3a na formação de placas vulneráveis e angiogênese de AS.
Materiais e métodos
Declaração de Ética
Os animais foram tratados com humanidade usando procedimentos aprovados em conformidade com as recomendações do Guia para o Cuidado e Uso de Animais de Laboratório do National Institutes of Health. O protocolo foi aprovado pelo Comitê de Uso e Cuidado Institucional de Animais do Hospital Popular da Província de Qinghai (número ético:201870726).
Animais Experimentais
ApoE masculina - / - camundongos e camundongos C57BL / 6J (grau livre de patógenos específicos) com idade de 8 semanas estavam disponíveis em Beijing Vital Laboratory Animal Technology (Pequim, China). Os camundongos (5-6 camundongos em uma gaiola) foram alojados em um ciclo de 12 h / 12 h dia / noite com acesso ad libitum a comida e água.
Estabelecimento de modelos de camundongos de AS
ApoE - / - camundongos foram alimentados com ração rica em gordura por 16 semanas para estabelecer o modelo de placa vulnerável AS. Camundongos C57BL / 6 J foram usados como o grupo normal com bebida e comida natural. ApoE - / - camundongos tinham cabelos claros e brilhantes e queda de cabelo nas costas após 12 semanas. O arco aórtico e a artéria braquiocefálica de 3 camundongos modelo foram dissecados para realizar a coloração com hematoxilina-eosina (HE), e não houve depósitos de placa significativos na íntima. Outros 3 camundongos modelados foram identificados novamente após 4 w, e a coloração HE mostrou que havia depósitos de placa óbvios na íntima do arco da aorta, indicando sucesso no estabelecimento do modelo.
Agrupamento e tratamento de camundongos
ApoE - / - camundongos com placa vulnerável a AS foram divididos em 6 grupos com 12 camundongos em cada grupo:grupo AS, grupo de controle negativo (NC) (injetado com solução salina normal em ApoE - / - camundongos), grupo miR-342-5p agomir (injetado com miR-342-5p agomir para superexpressar a expressão de miR-342-5p em ApoE - / - camundongos), grupo miR-342-5p antagomir (injetado com miR-342-5p antagomir para reduzir a expressão de miR-342-5p em ApoE - / - camundongos), grupo de superexpressão (oe) -Wnt3a (injetado com o vetor oe-Wnt3a para regular para cima a expressão de Wnt3a em ApoE - / - camundongos) e miR-342-5p agomir + grupo oe-Wnt3a (injetado com miR-342-5p agomir e vetor oe-Wnt3a para regular a expressão de miR-342-5p e Wnt3a em ApoE - / - camundongos). Camundongos C57BL / 6 J como o grupo normal foram alimentados com dieta normal. A ração rica em gordura continha 20% de gordura e 0,25% de colesterol. miR-342-5p agomir, miR-342-5p antagomir e o vetor oe-Wnt3a foram adquiridos da Sangon (Shanghai, China). O vetor oe-Wnt3a, miR-342-5p agomir, miR-342-5p antagomir foram todos dissolvidos em 0,2 mL de solução salina normal e injetados em camundongos a uma dose de 40 mg / kg através da veia da cauda a cada duas semanas. Após 8 semanas, amostras de sangue foram retiradas do globo ocular e, em seguida, os camundongos foram sacrificados para coletar tecidos arteriais [19].
No experimento preliminar, ApoE - / - camundongos com AS foram injetados com 10 mg / kg, 20 mg / kg, 40 mg / kg miR-342-5p agomir, miR-342-5p antagomir ou vetor oe-Wnt3a (uma vez a cada duas semanas; 4 vezes no total). Em seguida, os níveis de expressão de β-catenina foram detectados por reação em cadeia da polimerase quantitativa de transcrição reversa (RT-qPCR).
Coleta e tratamento de amostras
Antes da amostragem, os camundongos jejuaram por 12 horas e foram anestesiados por inalação de éter e amostras de sangue foram coletadas dos globos oculares. O tórax dos camundongos foi aberto, a aorta torácica foi dissociada ao final da aorta abdominal e todo o vaso foi removido. Depois de limpos por solução salina tamponada com fosfato livre de RNA (PBS), os tecidos foram incluídos para coloração HE, óleo vermelho O, coloração com vermelho Sirius e coloração imuno-histoquímica. Alguns dos tecidos vasculares foram preservados a -80 ° C para RT-qPCR e Western blot.
Detecção do nível de lipídios no sangue
O analisador bioquímico automático (Roche, Basel, Suíça) foi adotado para detectar colesterol total (TC), triglicerídeo (TG), colesterol de lipoproteína de baixa densidade (LDL-C) e colesterol de lipoproteína de alta densidade (HDL-C) no soro. A detecção foi implementada seguindo as especificações dos kits (NanJing JianCheng Bioengineering Institute, Nanjing, China).
Ensaio de imunoabsorção enzimática (ELISA)
Determinação do conteúdo de citocinas séricas:foram usados kits comerciais de interleucina (IL) -5, IL-12p70, fator de necrose tumoral alfa (TNF-α) e interferon (IFN) -γ ELISA. Finalmente, o valor de densidade óptica (DO) de cada poço foi testado por um leitor de microplaca a 450 nm.
Determinação da lesão por estresse oxidativo:o conteúdo de malondialdeído (MDA) e a atividade da superóxido dismutase (SOD) no soro foram testados pelo kit MDA (o valor OD foi testado por espectrofotômetro a 532 nm) e o kit SOD (o valor OD foi determinado por um leitor de microplaca a 450 nm). Os kits IL-5, IL-12p70, TNF-α, IFN-γ, MDA e SOD ELISA foram adquiridos na MultiSciences (Lianke) Biotechnology Corporate Limited (Hangzhou, Zhejiang, China).
Coloração HE, Coloração Oil Red O e Coloração Sirius Red
Após a fixação e incorporação, os espécimes foram fatiados em seções consecutivas de 4 microm de espessura. As fatias foram desparafinadas e hidratadas, coradas com hematoxilina e eosina, diferenciadas, desidratadas, depuradas com xilol, secas e seladas com goma neutra. O núcleo era azul e outros tecidos, como citoplasma e tecidos conjuntivos, eram vermelhos em tons diferentes. A formação da placa foi observada por um microscópio de fluorescência. As paredes arteriais dos cortes corados com HE foram selecionadas ao microscópio, e os resultados experimentais foram coletados por uma câmera digital. O módulo de software de análise de imagem Image Pro Plus6.0 (IPP6.0) foi utilizado para calcular a área da placa da seção transversal de cada corte e a área da parede, bem como sua proporção.
Fatias com 4-5 μm foram escolhidas para a coloração com óleo vermelho O. As fatias foram secas com excesso de temperatura por 20 min e incubadas com isopropanol 100% por 5 min. Em seguida, as fatias foram incubadas com solução de coloração de óleo vermelho O 0,5% em um forno a 60 ° C por 8 min, lavadas em isopropanol 85% por 3 min, tingidas com hematoxilina por 1 min, limpas e seladas. Os resultados da coloração com óleo vermelho O sugeriram que o lipídio era vermelho ou laranja e o núcleo era azul claro. O software IPP6.0 foi usado para calcular a área de gordura e a área da placa na placa da fatia de tecido. O teor de lípidos =área de coloração positiva com vermelho de óleo O / área da placa × 100%.
Coloração com vermelho Sirius:as fatias foram desparafinadas e hidratadas, tingidas por 10 minutos com solução de coloração com azul celestino, com solução de coloração com vermelho Sirius por 20 minutos e contrastadas por 10 minutos com hematoxilina. Por fim, as fatias foram desidratadas em gradiente de etanol, clarificadas com xilol e seladas com goma neutra. A área de colágeno na placa da fatia de tecido foi calculada pelo software IPP6.0. A área de colágeno =área de coloração positiva de Sirius red / área de placa × 100%. Calculou-se a porcentagem de lipídio e colágeno na área da placa.
Hematoxilina, eosina e corante Sirius estavam disponíveis no China Pharmaceutical Group Shanghai Chemical Reagent Co. Ltd. (Xangai, China). O óleo vermelho O em pó foi adquirido à Sigma-Aldrich Chemical Company (St Louis, MO, EUA).
Reação em cadeia da polimerase quantitativa com transcrição reversa (RT-qPCR)
Os tecidos aórticos foram adicionados ao reagente de extração de RNA total Trizol (Invitrogen, Carlsbad, Califórnia, EUA) e a seguir homogeneizados para extrair o RNA total e o DNA complementar composto. Os primers foram todos referenciados à sequência fornecida pelo Genbank, projetado pelo Primer 5.0 e sintetizado pela Shanghai Sangon Biotechnology Co. Ltd. MiR-342-5p:forward:5′-CGGAGGGGTGCTATCTGTGATTGAG-3 ′, os primers reversos foram kits universais de primers (Qiagen empresa, Hilden, Alemanha); Wnt3a:direto:5′-AGGTAAGCTACTCCCTCAACTA-3 ′, reverso:5′-CTGAAGCACCCTCTCATGTATC-3 ′; β-actina:direto:5′-GCACCACACCTTCTACAATGAGC -3 ′, reverso:5′-TCGTTGCCAATAGTGATGACC-3 ′; β-catenina:direto:5′-TCAAGAGAGCAAGCTCATCATTCT-3 ′, reverso:5′-CACCTTCAGCACTCTGCTTGTG-3 ′. Após a reação, o ciclo de limiar (Ct) foi analisado por computador. A razão relativa de miR-342-5p para U6 foi usada como sua expressão, a razão relativa de Wnt3a para β-actina foi usada como sua expressão e a razão relativa foi calculada por 2 −ΔΔCt método.
Análise de Western Blot
A proteína total foi extraída dos tecidos aórticos. A concentração de proteína foi medida pelo método do ácido bicinconínico. Foi realizada eletroforese em gel de poliacrilamida. Em seguida, a proteína foi transferida para a membrana de fluoreto de polivinilideno e a banda alvo foi obtida. A membrana foi selada em 5% de leite desnatado por 1 h, adicionado com anticorpos primários Wnt3a (1:500), β-catenina (1:1000, Santa Cruz Biotechnology, Inc., Santa Cruz, CA, EUA), CD34 (1 :2500, Abcam, MA, EUA) e β-actina (1:2000, Beyotime Biotechnology Co., Xangai, China) a 4 ° C durante a noite. A membrana foi lavada por solução salina tamponada com Tris com Tween 20 (pH =7,5, Tris-HCl 10 mmol / L, NaCl 100 mmol / L e Tween-20 0,2%) por 10 min × 3 vezes e, em seguida, anexada com anticorpo secundário (1:1000, ZSGB-Bio, Pequim, China) por 2 h. O software ImageJ foi adotado para avaliar o valor de cinza das bandas e quantificar a expressão da proteína.
Coloração imunohistoquímica
Fatias de 4–5 µm foram colocadas nas lâminas revestidas com polilisina 100 mg / L e fixadas com acetona. A peroxidase endógena foi bloqueada pela albumina de soro bovino. Os tecidos foram gotejados com anticorpo MOMA-2 (1:200), α-SMA (1:200) e CD34 (1:200, Abcam Inc., Cambridge, MA, EUA), bem como adicionado com solução de trabalho de anticorpo secundário ( 1:1000). Os tecidos foram revelados por diaminobenzidina, contrastados por hematoxilina (1 min), desidratados, clarificados, selados e observados ao microscópio. Três campos visuais diferentes foram selecionados para cada seção de imunohistoquímica. O software IPP6.0 foi executado para análise quantitativa. A coloração imuno-histoquímica positiva de MOMA-2 e α-SMA, respectivamente, indica que macrófagos e células de músculo liso estão localizados principalmente no citoplasma, que é amarelo a marrom. As porcentagens de macrófagos e células musculares lisas foram calculadas separadamente, as quais foram combinadas com a porcentagem de lipídio e colágeno na placa para calcular o índice de vulnerabilidade da placa. O índice de vulnerabilidade da placa =(porcentagem positiva de macrófagos + porcentagem positiva de lipídios) / (porcentagem positiva de colágeno + porcentagem positiva de células musculares lisas) [20]. A densidade dos microvasos (MVD) foi avaliada pela medição da expressão de CD34 e quantificada como o número de microvasos / mm 2 .
Ensaio do gene Dual Luciferase Reporter
O gene alvo do miR-342-5p foi analisado pelo site de predição biológica (http://www.microRNA.org). O ensaio do gene repórter de luciferase dupla foi usado para verificar se Wnt3a era o gene alvo de miR-342-5p. A sequência de tipo selvagem ou mutante da região 3'-não traduzida de Wnt3a (3'-UTR) foi clonada no vetor GP-miRGLO (GenePharma, Shanghai, China). O repórter (0,5 μg) e 1, 10 ou 100 pM miR-342-5p agomir foram transfectados em células endoteliais aórticas de camundongo (No. 506, MingzhouBio, Ningbo, China) por 48 h para testar a atividade da luciferase usando o sistema de ensaio de luciferase dupla (Promega, WI, EUA).
Análise estatística
Todos os dados foram interpretados pelo software SPSS 21.0 (IBM Corp. Armonk, NY, EUA). Os dados de medição foram indicados como média ± desvio padrão. Disparidades entre dois grupos foram formuladas por t -teste, enquanto aqueles entre vários grupos por análise de variância unilateral (ANOVA) seguida pelo teste de comparações múltiplas de Tukey. A significância estatística foi estabelecida por P valor <0,05.
Resultados
Aumentos de miR-342-5p e diminuições de Wnt3a nos tecidos da aorta de ApoE - / - ratos e miR-342-5p alveja diretamente Wnt3a
Os genes alvo de microRNA (miRNA) estão associados a funções relacionadas à aterosclerose. miR-342-5p, Wnt3a e β-catenina foram testados em tecidos aórticos de camundongos modelo AS por RT-qPCR e ensaio de Western blot. Foi revelado que em relação ao grupo normal, miR-342-5p foi aumentado enquanto Wnt3a e β-catenina diminuíram no grupo AS (ambos P <0,05). Em comparação com o grupo NC, miR-342-5p foi aprimorado, bem como Wnt3a e β-catenina foram diminuídos no grupo agomir miR-342-5p (ambos P <0,05), enquanto miR-342-5p foi diminuído, Wnt3a e β-catenina foram aumentados no grupo de antagomir miR-342-5p (ambos P <0,05). A expressão de Wnt3a e β-catenina foi aumentada no grupo oe-Wnt3a em relação ao grupo NC (ambos P <0,05). Em comparação com o grupo miR-342-5p agomir, a expressão de Wnt3a e β-catenina aumentou no grupo miR-342-5p agomir + oe-Wnt3a ( P <0,05) (Fig. 1A – D). Além disso, no experimento preliminar, a expressão de β-catenina sob o tratamento de diferentes concentrações de miR-342-5p agomir, miR-342-5p antagomir e oe-Wnt3a foi testada e os resultados apresentados (Arquivo adicional 1:Fig. S1 ) quanto maior a concentração de miR-342-5p agomir, menor a expressão de β-catenina; quanto maior a concentração de antagomir miR-342-5p, maior a expressão de β-catenina; e quanto maior a concentração de oe-Wnt3a, maior a expressão de β-catenina.
miR-342-5p aumenta e Wnt3a diminui em tecidos aórticos de ApoE - / - camundongos e miR-342-5p tem como alvo direto Wnt3a. A Expressão de miR-342-5p em tecido aórtico de camundongos detectado por RT-qPCR. B Expressão de mRNA de Wnt3a em tecido aórtico de camundongos detectado por RT-qPCR ( n =12). C , D Expressão da proteína Wnt3a e β-catenina em tecidos aórticos de camundongos testados por análise de Western blot ( n =12). E Local de ligação de miR-342-5p em Wnt3a 3′-UTR. F miR-342-5p agomir diminuiu de forma dependente da dose a atividade relativa nas células transfectadas com Wnt3a 3′-UTR ( N =3). G Atividade relativa da luciferase em células com tipo selvagem e mutante Wnt3a 3′-UTR após transfectada com miR-342-5p agomir ou scramble ( N =3). * P <0,05 vs. o grupo normal, # P <0,05 vs. o grupo NC. & P <0,05 vs. o grupo miR-342-5p agomir. Os dados de medição foram indicados como média ± desvio padrão. As comparações entre dois grupos foram formuladas por t -teste, enquanto as comparações entre vários grupos foram avaliadas por ANOVA de uma via seguida pelo teste de comparações múltiplas de Tukey. AS, aterosclerose; NC, controle negativo
Os miRNAs podem inibir a tradução de genes específicos ligando-se ao seu RNA mensageiro 3′UTR. O site de bioinformática previu que havia uma relação de destino entre miR-342-5p e Wnt3a (Fig. 1E). O ensaio do gene repórter de luciferase dupla relatou que em células endoteliais aórticas de camundongos transfectadas com vetor Wnt3a 3′UTR, o valor de renilla / vaga-lume da luciferase foi reduzido de forma dependente da dose por transfecção com miR-342-5p agomir, com uma diminuição significativa de 10 para 100 pM miR-342-5p agomir e uma diminuição de 64% ocorreu no grupo 100 pM miR-342-5p agomir quando comparado com o grupo NC. Isso indicou a presença de um local alvo miR-342-5p no Wnt3a 3′UTR. No entanto, o valor renilla / firefly da atividade da luciferase não foi afetado no grupo de mutação Wnt3a (Fig. 1F, G). Assim, pode ser confirmado que Wnt3a era um gene alvo direto de miR-342-5p, e miR-342-5p / Wnt3a poderia regular a progressão de AS.
Efeitos de Wnt3a regulado para cima ou miR-342-5p regulado para baixo nos níveis de lipídios em ApoE - / - Ratos
Além disso, para investigar se miR-342-5p direcionar e regular a via de sinalização Wnt3a afetaria os níveis de lipídios de camundongos AS, analisador bioquímico automático foi utilizado para observar a mudança dos níveis de lipídios. Os resultados revelaram que (Fig. 2A-D) em contraste com o grupo normal, os conteúdos de TC, TG e LDL-C foram aumentados e o conteúdo de HDL-C diminuiu no grupo AS (todos P <0,05). Contra o grupo NC, os conteúdos de TC, TG e LDL-C foram aumentados e o conteúdo de HDL-C diminuiu no grupo agomir miR-342-5p (todos P <0,05), enquanto os conteúdos de TC, TG e LDL-C diminuíram e o conteúdo de HDL-C aumentou no grupo de antagomir miR-342-5p e no grupo oe-Wnt3a (todos P <0,05). Em relação ao grupo miR-342-5p agomir, os conteúdos de TC, TG e LDL-C foram reduzidos e o conteúdo de HDL-C aumentou no grupo miR-342-5p agomir + oe-Wnt3a (todos P <0,05). Estes resultados sugerem que miR-342-5p e Wnt3a regulam o nível de lipídios no sangue de camundongos AS e ilustram ainda mais a relação de regulação direcionada entre miR-342-5p e Wnt3a. A superexpressão de Wnt3a reverteria os efeitos induzidos por miR-342-5p superexpressado em camundongos AS.
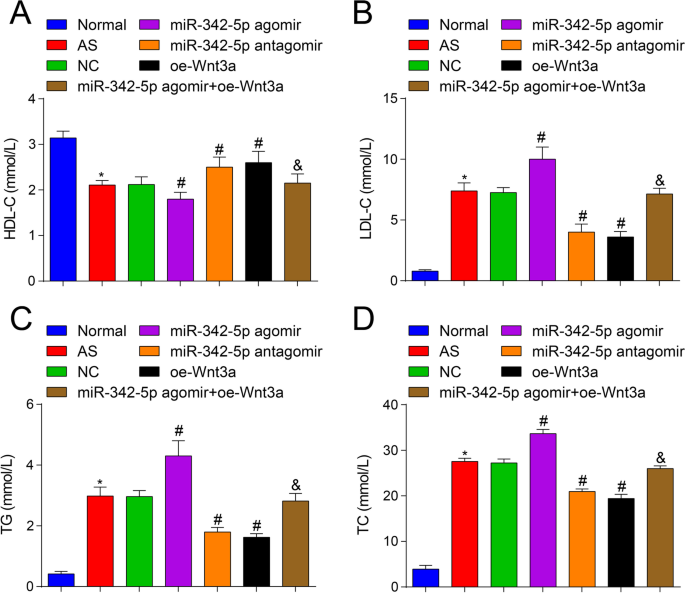
Efeitos de Wnt3a regulado para cima ou miR-342-5p regulado para baixo nos níveis de lipídios em ApoE - / - camundongos. A Comparação do conteúdo de HDL-C no soro do grupo de camundongos. B Comparação dos conteúdos de LDL-C no soro do grupo de camundongos. C Comparação dos conteúdos de TG no soro do grupo de camundongos. D Comparação dos conteúdos de TC no soro do grupo de camundongos. n =12. * P <0,05 vs. o grupo normal, # P <0,05 vs. o grupo NC. & P <0,05 vs. o grupo miR-342-5p agomir. Os dados de medição foram indicados como média ± desvio padrão. As comparações entre vários grupos foram avaliadas por ANOVA de uma via seguida pelo teste de comparações múltiplas de Tukey. AS, aterosclerose; NC, controle negativo
Efeitos da superexpressão de Wnt3a ou baixa expressão de miR-342-5p em citocinas inflamatórias e oxidativas relacionadas ao estresse no soro de ApoE - / - Ratos
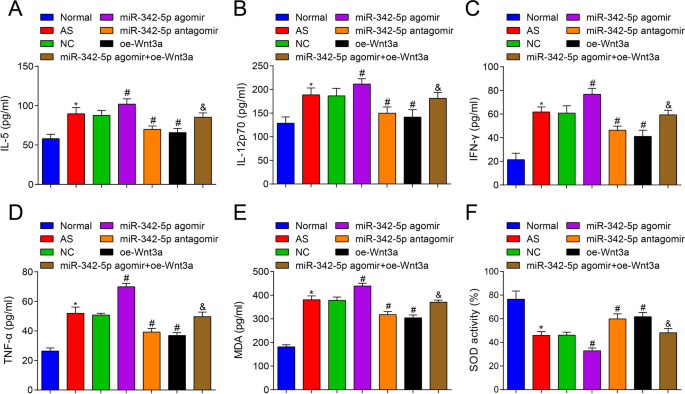
Em seguida, o conteúdo de citocinas no soro de camundongos AS foi testado por ELISA, e os resultados relataram que (Fig. 3A-D) versus o grupo normal, IL-5, IL-12p70, IFN-γ e TNF-α foram aumentados no grupo AS (todos P <0,05). Contra o grupo NC, os conteúdos de IL-5, IL-12p70, IFN-γ e TNF-α foram aumentados no grupo agomir miR-342-5p (todos P <0,05), enquanto os conteúdos de IL-5, IL-12p70, IFN-γ e TNF-α foram degradados no grupo antagomir miR-342-5p e no grupo oe-Wnt3a (todos P <0,05). Em contraste com o grupo miR-342-5p agomir, os conteúdos de IL-5, IL-12p70, IFN-γ e TNF-α foram diminuídos no grupo miR-342-5p agomir + oe-Wnt3a (todos P <0,05). Foi sugerido que a regulação direcionada do miR-342-5p da via de sinalização Wnt3a regulou ainda mais o nível de citocinas relacionadas no soro de camundongos AS.
Efeitos da superexpressão de Wnt3a ou baixa expressão de miR-342-5p em citocinas relacionadas ao estresse oxidativo e inflamatório no soro de ApoE - / - camundongos. A Comparação dos conteúdos de IL-5 no soro do grupo de camundongos. B Comparação dos conteúdos de IL-12p70 no soro do grupo de camundongos. C Comparação dos conteúdos de IFN-γ no soro do grupo de camundongos. D Comparação dos conteúdos de TNF-α no soro do grupo de camundongos. E, Comparação do conteúdo de MDA no soro do grupo de camundongos. F Comparação da atividade SOD no soro do grupo de camundongos. n =12. * P <0,05 vs. o grupo normal, # P <0,05 vs. o grupo NC. & P <0,05 vs. o grupo miR-342-5p agomir. Os dados de medição foram indicados como média ± desvio padrão. As comparações entre vários grupos foram avaliadas por ANOVA de uma via seguida pelo teste de comparações múltiplas de Tukey. AS, aterosclerose; NC, controle negativo
Além disso, o conteúdo de MDA e a atividade de SOD no soro de camundongos foram testados e foi revelado que (Fig. 3E, F) em comparação com o grupo normal, o conteúdo de MDA foi aumentado e a atividade de SOD foi reduzida no grupo de AS (ambos P <0,05). Em comparação com o grupo NC, o conteúdo de MDA foi aumentado e a atividade de SOD diminuiu no grupo agomir miR-342-5p (ambos P <0,05), enquanto o conteúdo de MDA diminuiu e a atividade de SOD aumentou no grupo miR-342-5p antagomir e no grupo oe-Wnt3a (todos P <0,05). Contra o grupo agomir miR-342-5p, o conteúdo de MDA foi deprimido e a atividade de SOD foi aumentada no grupo miR-342-5p agomir + oe-Wnt3a (ambos P <0,05). Portanto, foi obtido um resumo de que a depleção de miR-342-5p e a restauração de Wnt3a inibiram o estresse oxidativo em camundongos AS.
Efeitos do esgotamento de miR-342-5p ou restauração de Wnt3a no conteúdo de lipídios e colágeno na placa aórtica de ApoE - / - Ratos
A fim de explorar o efeito de miR-342-5p direcionado a Wnt3a na área da placa em tecido aórtico de camundongos, a coloração HE foi realizada e os resultados revelaram que (Fig. 4A, B) exceto para o grupo normal, placas AS formadas em todos seções dos outros grupos. No grupo AS, a área da placa era grande, a capa fibrosa era mais fina, o núcleo lipídico estava aumentado, mais células de espuma e precipitação de cristais de colesterol apareceram na placa, a parede interna da artéria e a camada muscular estavam espessas e a placa estava instável. A situação do grupo NC era semelhante à do grupo AS. No grupo miR-342-5p antagomir e no grupo oe-Wnt3a, a área da placa era pequena, a íntima da artéria era lisa e as capas fibrosas tinham número reduzido e tornaram-se mais finas. Não houve ruptura, mas sim células espumosas de diferentes tamanhos na placa. O cristal de colesterol estava assimetricamente distribuído e parcialmente calcificado, o número de células musculares lisas e fibras de colágeno aumentaram e a placa tendeu a ser estável. Em comparação com o grupo NC, a área da placa foi aumentada e as lesões AS foram agravadas no grupo agomir miR-342-5p ( P <0,05) enquanto a área da placa diminuiu no grupo de antagomir miR-342-5p e no grupo oe-Wnt3a com lesões AS reduzidas (ambos P <0,05). Em comparação com o grupo miR-342-5p agomir, a área da placa foi diminuída no grupo miR-342-5p agomir + oe-Wnt3a ( P <0,05).
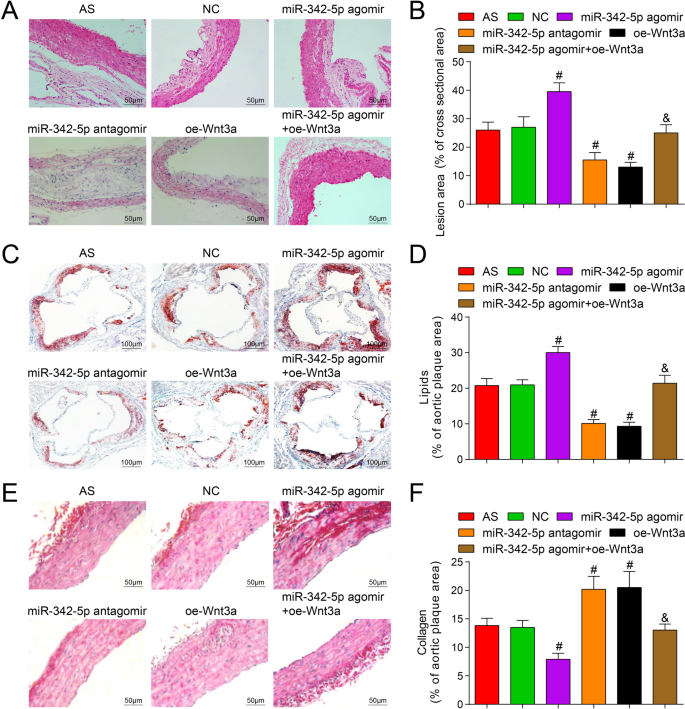
Efeitos da depleção de miR-342-5p ou restauração de Wnt3a no conteúdo de lipídios e colágeno na placa aórtica de ApoE - / - camundongos. A Resultados da coloração de HE aórtica em camundongos (barra de escala:50 μm). B Comparação da área da placa aórtica em cada grupo de camundongos. C Resultados da coloração aórtica com óleo vermelho O em todos os grupos de camundongos (barra de escala:100 μm). D Comparação do conteúdo lipídico no tecido aórtico de camundongos. E Resultados da coloração aórtica com vermelho Sirius em cada grupo de camundongos (barra de escala:50 μm). F Comparação do conteúdo de colágeno em tecidos aórticos de camundongos. n =12. # P <0,05 vs. o grupo NC. & P <0,05 vs. o grupo miR-342-5p agomir. Os dados de medição foram indicados como média ± desvio padrão. As comparações entre vários grupos foram avaliadas por ANOVA de uma via seguida pelo teste de comparações múltiplas de Tukey. AS, aterosclerose; NC, controle negativo
A coloração com óleo vermelho O e a coloração com vermelho Sirius foram adotadas para detectar o efeito de miR-342-5p direcionado Wnt3a no conteúdo de lipídios e conteúdo de colágeno na placa de tecido aórtico de camundongos, e os resultados demonstraram que (Fig. 4C-F) Vermelho de óleo A coloração O mostrou gordura vermelha e núcleo azul, enquanto a coloração com vermelho Sirius mostrou fibras de colágeno vermelhas e núcleo azul. Em comparação com o grupo NC, o conteúdo de lipídios foi aumentado e o conteúdo de colágeno foi reduzido no grupo miR-342-5p agomir, bem como o conteúdo lipídico foi reduzido e o conteúdo de colágeno foi acumulado no grupo antagomir miR-342-5p e oe-Wnt3a grupo (todos P <0,05). Com relação ao grupo miR-342-5p agomir, o conteúdo de lipídios foi diminuído e o conteúdo de colágeno foi elevado no grupo miR-342-5p agomir + oe-Wnt3a (ambos P <0,05). Os resultados experimentais ilustraram totalmente que a regulação direcionada do miR-342-5p da via de sinalização Wnt3a teve um efeito regulatório sobre o conteúdo de lipídios e colágeno na placa aórtica de camundongos AS.
Efeitos de miR-342-5p regulado para baixo ou Wnt3a regulado para cima em macrófagos e células musculares lisas na placa aórtica de ApoE - / - Ratos
O grau de AS é diretamente proporcional ao conteúdo de macrófagos mononucleares [21]. VSMCs são as células principais na camada média das artérias e são essenciais para manter a integridade da parede arterial. VSMCs estão envolvidos na reconstrução da parede arterial e desempenham um papel importante na EA em vários estágios [22]. α-SMA é um marcador específico de células musculares lisas [23]. Neste estudo, um anticorpo marcador de macrófago (MOMA-2) foi utilizado para marcar macrófagos, e imunohistoquímica foi aplicada para detectar a expressão de MOMA-2 e α-SMA, respectivamente.
Ao microscópio, a coloração imuno-histoquímica positiva de MOMA-2 e α-SMA, respectivamente, indica que macrófagos e células musculares lisas estão localizados principalmente no citoplasma, que é amarelo a marrom. O MOMA-2 imuno positivo indicou que os macrófagos estavam localizados principalmente no citoplasma com amarelo a marrom. Determinado por imunohistoquímica, foi manifestado que versus o grupo NC, a porcentagem de coloração positiva de macrófagos de placa (MAMO-2) aumentou e a porcentagem de células de músculo liso positivas diminuiu no agomir miR-342-5p (ambos P <0,05). A porcentagem de coloração positiva de macrófagos de placa (MAMO-2) foi reduzida e a porcentagem de células de músculo liso positivas foi aumentada no grupo miR-342-5p antagomir e no grupo oe-Wnt3a (todos P <0,05). Em comparação com o grupo miR-342-5p agomir, a porcentagem de macrófagos da placa (MAMO-2) coloração positiva foi diminuída e a porcentagem de células de músculo liso positivas aumentou no grupo miR-342-5p agomir + oe-Wnt3a (ambos P <0,05) (Fig. 5A-D). Ficou implícito que a regulação direcionada do miR-342-5p da via de sinalização Wnt3a poderia regular a agregação de macrófagos e células do músculo liso em placas de tecido arterial de camundongos AS.
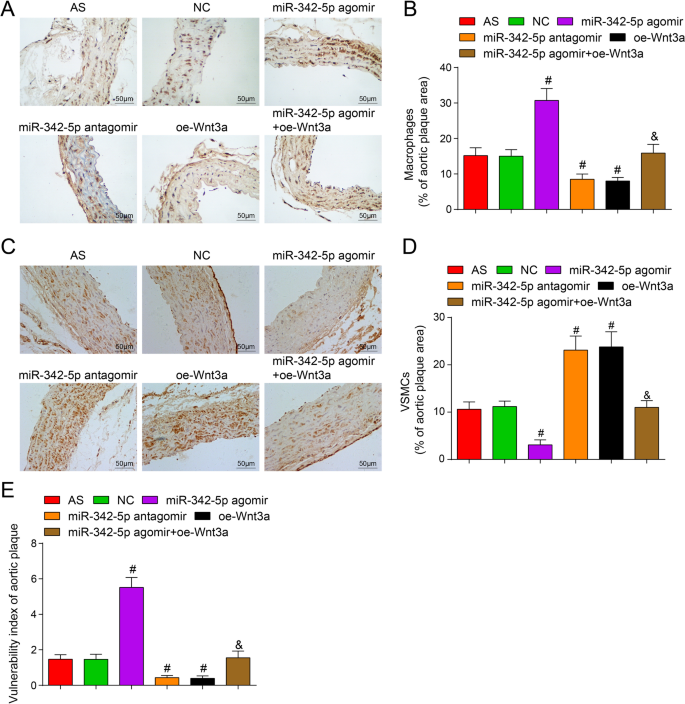
Efeitos da alta expressão de Wnt3a ou má expressão de miR-342-5p na vulnerabilidade da placa aórtica de ApoE - / - camundongos. A Coloração imunohistoquímica de MOMA-2 em cada grupo de camundongos (barra de escala:50 μm). B Análise quantitativa da figura A. C Coloração imunohistoquímica de α-SMA em cada grupo de camundongos (barra de escala:50 μm). D Análise quantitativa da figura C . E Comparação do índice de vulnerabilidade da placa em tecidos aórticos de camundongos AS. n =12. # P <0,05 vs. o grupo NC. & P <0,05 vs. o grupo miR-342-5p agomir. Os dados de medição foram indicados como média ± desvio padrão. As comparações entre vários grupos foram avaliadas por ANOVA de uma via seguida pelo teste de comparações múltiplas de Tukey. AS, aterosclerose; NC, controle negativo
Efeitos da alta expressão de Wnt3a ou má expressão de miR-342-5p na vulnerabilidade da placa aórtica de ApoE - / - Ratos
O índice de vulnerabilidade da placa foi calculado:(porcentagem positiva de macrófagos + porcentagem positiva de lipídios) / (porcentagem positiva de células musculares lisas + porcentagem positiva de colágeno). Em relação ao grupo NC, o índice de vulnerabilidade de placa foi elevado no grupo miR-342-5p agomir ( P <0,05) e diminuiu no grupo de antagomir miR-342-5p e no grupo oe-Wnt3a (ambos P <0,05). Em comparação com o grupo miR-342-5p agomir, o índice de vulnerabilidade de placa foi diminuído no grupo miR-342-5p agomir + oe-Wnt3a ( P <0,05) (Fig. 5E). Resumidamente, miR-342-5p direcionou a regulação da vulnerabilidade mediada pela via de sinalização Wnt3a de placas em tecidos arteriais de camundongos AS.
Efeitos da baixa expressão de miR-342-5p ou superexpressão de Wnt3a na angiogênese na placa aórtica de ApoE - / - Ratos
Os anticorpos contra o marcador de células endoteliais CD34 podem detectar a densidade dos vasos sanguíneos [24]. Por imunohistoquímica e Western blot. Em comparação com o grupo NC, o MVD foi aumentado no grupo miR-342-5p agomir e atenuado no grupo miR-342-5p antagomir e no grupo oe-Wnt3a (todos P <0,05). Em comparação com o grupo miR-342-5p agomir, o MVD foi diminuído no grupo miR-342-5p agomir + oe-Wnt3a ( P <0,05) (Fig. 6A-C). Coletivamente, miR-342-5p direcionando e regulando a via de sinalização Wnt3a participou diretamente na regulação da densidade de neovascularização em placas de camundongos AS.
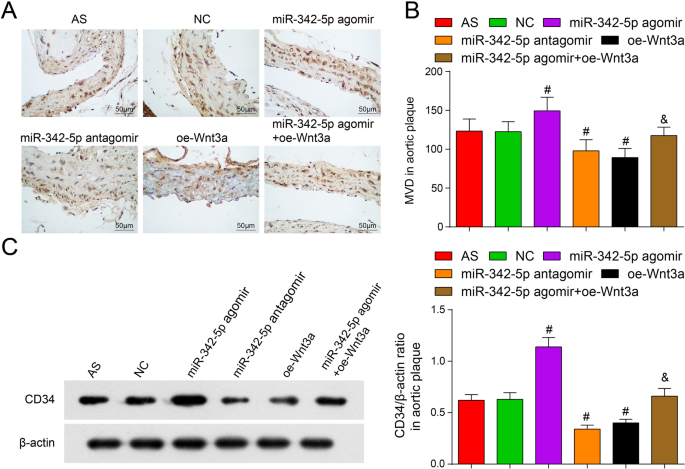
Efeitos da baixa expressão de miR-342-5p ou superexpressão de Wnt3a em MVD na placa aórtica de ApoE - / - camundongos. A Coloração imunohistoquímica de CD34 em cada grupo de ApoE - / - camundongos (barra de escala:50 μm). B Comparação de MVD em placa aórtica em ApoE - / - camundongos. C Comparação da expressão da proteína CD34 em ApoE - / - camundongos. n =12. # P <0,05 vs. o grupo NC. & P <0,05 vs. o grupo miR-342-5p agomir. Os dados de medição foram indicados como média ± desvio padrão. As comparações entre vários grupos foram avaliadas por ANOVA de uma via seguida pelo teste de comparações múltiplas de Tukey. AS, aterosclerose; NC, controle negativo
Discussão
AS é uma doença imprevisível que envolve formas de inflamação crônica e remodelação vascular, e é a principal causa de mortalidade e morbidade em todo o mundo [25]. Um estudo anterior discutiu que miR-342-5p está implicado na regulação da progressão da AS [26]. Além disso, foi refletido que a via Wnt está envolvida em facilitar a ocorrência e o desenvolvimento de EA diabético [27]. Como os mecanismos relacionados de miR-342-5p e Wnt3a em AS ainda precisam ser escavados, nosso estudo foi investigar o efeito de miR-342-5p direcionado a Wnt3a na formação de placa vulnerável e angiogênese de AS.
Nosso estudo revelou que miR-342-5p altamente expresso e Wnt3a pouco expresso foram encontrados em tecidos aórticos de camundongos AS. Um estudo apresentou que miR-342-5p derivado de macrófago é dramaticamente aumentado em lesões ateroscleróticas iniciais em ApoE - / - ratos [13]. Outro estudo apresentou que miR-342-5p é marcadamente elevado em pacientes com fibrilação atrial [28]. Foi relatado que a deficiência de Wnt3a prejudica irreversivelmente a autorrenovação das células-tronco hematopoéticas e resulta em defeitos na diferenciação das células progenitoras [29]. Um estudo mostrou que a depleção de Wnt3a leva a função cardíaca defeituosa [30]. Foi revelado que a expressão de Wnt3a no hipocampo de camundongos com doença de Alzheimer está notavelmente diminuída [31]. Outro resultado de nosso estudo é que Wnt3a foi diretamente direcionado por miR-342-5p em camundongos AS. Foi relatado que miR-342-5p pode ter como alvo a 3'-UTR de Wnt3a e regular negativamente sua expressão [14].
Além disso, nosso estudo sugeriu que os conteúdos de TC, TG, LDL-C, IL-5, IL-12p70, IFN-γ, TNF-α e MDA estavam aumentados no soro e o conteúdo de HDL-C e a atividade de SOD diminuíram. Além disso, a área da placa, o conteúdo de lipídios, o conteúdo de colágeno e o MVD foram aumentados, bem como a expressão de MOMA-2 foi aumentada e a expressão de α-SMA foi diminuída em camundongos AS. O IFN-γ é uma citocina solúvel com muitas funções, incluindo antifibrose, antiproliferação, imunomodulação, apoptose e atividades antivirais [32]. Foi revelado que o tratamento com glutamina aumenta significativamente a atividade de SOD e reduz o conteúdo de MDA, bem como aumenta os níveis de proteína Wnt3a na doença de Alzheimer [31]. Um estudo revelou que os níveis plasmáticos de TC, TG e LDL-C estão notavelmente elevados e o HDL-C está acentuadamente reduzido na EA [33]. Um estudo relatou que a suplementação de TEMPOL, que tem valor na supressão de distúrbios metabólicos e no aumento da estabilidade da placa aterosclerótica, aumenta o conteúdo de colágeno da placa e reduz o conteúdo de lipídios [34]. Zhou et al. observou que os tratamentos OPCRR reduzem drasticamente os perfis lipídicos séricos, incluindo TC, TG e LDL-C, bem como aumenta o HDL-C, também diminui o conteúdo de MDA como um produto da peroxidação lipídica e, além disso, diminui os níveis séricos de TNF-α em AS [35]. Foi apresentado que as amostras ateroscleróticas obviamente reduziram a expressão de α-SMA [36]. Um estudo apresentou que o MVD elevado é encontrado em aortas doentes e, especialmente, na ruptura da placa aterosclerótica [37]. Além disso, nosso estudo revelou que a expressão pobre de miR-342-5p e a superexpressão de Wnt3a diminuíram os níveis de lipídios, o conteúdo de citocinas, a resposta ao estresse oxidativo, a área da placa e o conteúdo lipídico, bem como o aumento do conteúdo de colágeno, a expressão de MOMA-2 depletada e o α restaurado -Expressão de SMA em tecidos aórticos em camundongos AS. Foi sugerido anteriormente que miR-342-5p está positivamente ligado aos níveis séricos de LDL-C e TNF-α e tem uma correlação inversa com HDL-C em pacientes com doença arterial coronariana (DAC) [12]. Outro estudo verificou que a depleção de miR-342-5p inibe AS [13]. Além disso, um experimento apresentou que o baixo nível sérico de Wnt1 está relacionado ao aumento de TG e LDL-C em paciente com DAC prematuro [38]. Além disso, um estudo mostrou que Wnt3a regulado positivamente, conteúdo aumentado de SOD e conteúdo diminuído de MDA são encontrados nos grupos de curcumina em ratos com doença de Parkinson [39].
Conclusão
Em resumo, nosso estudo pela primeira vez descobriu o mecanismo do eixo miR-342-5p / Wnt3 em AS e revelou que o esgotamento de miR-342-5p poderia reduzir a formação de placa vulnerável e angiogênese em camundongos AS por meio da restauração de Wnt3a, que pode ser um candidato potencial para o tratamento de AS (arquivo adicional 2:Fig. S2). O miR-342-5p pode ter um efeito sinérgico com outros miRNAs na doença vascular aterosclerótica, mas devido às restrições de tempo e financiamento, não conduzimos discussões relevantes adicionais, o que também é uma limitação deste estudo.
Disponibilidade de dados e materiais
As contribuições originais apresentadas no estudo estão incluídas no artigo / Material Suplementar, e outras consultas podem ser dirigidas ao autor para correspondência.
Abreviações
- miR-342-5p:
-
MicroRNA-342-5p
- A:
-
Aterosclerose
- α-SMA:
-
α-Actina de músculo liso
- MVD:
-
Densidade de microvasos
- miRNA:
-
MicroRNA
- oe:
-
Superexpressão
- NC:
-
Controle negativo
- PBS:
-
Salina tamponada com fosfato
- TC:
-
Colesterol total
- TG:
-
Triglicerídeo
- LDL-C:
-
Colesterol de lipoproteína de baixa densidade
- HDL-C:
-
Colesterol de lipoproteína de alta densidade
- ELISA:
-
Ensaio de imunoabsorção enzimática
- IL:
-
Interleucina
- TNF-α:
-
Fator de necrose tumoral alfa
- IFN:
-
Interferon
- MDA:
-
Malondialdeyde
- SOD:
-
Superoxido dismutação
- RT-qPCR:
-
Reação em cadeia da polimerase quantitativa com transcrição reversa
Avanços e desafios dos nanomateriais fluorescentes para aplicações biomédicas e de síntese
Tecnologias de Deposição de Camada Atômica Avançada para Micro-LEDs e VCSELs
Nanomateriais
- Caracterizando desvios de frequência de cristais de quartzo:tolerância de frequência, estabilidade de frequência e envelhecimento
- O monitoramento das condições aumenta e mantém a eficiência na produção de papel
- Criação e geração de imagens do Ciclocarbon
- Grafeno em alto-falantes e fones de ouvido
- A regulação negativa de microRNA-342-5p ou a regulação positiva de Wnt3a inibe a angiogênese e mantém a estabilidade da placa aterosclerótica em camundongos com aterosclerose
- Pesquisa Experimental sobre Estabilidade e Convecção Natural de Nanofluido de TiO2-Água em Compartimentos com Diferentes Ângulos de Rotação
- Supercapacitores elásticos miniaturizados e lineares de alta taxa
- Estudo de primeiros princípios sobre a estabilidade e imagem STM de Borophene
- Toxicidade potencial para fígado, cérebro e embrião de nanopartículas de dióxido de titânio em camundongos
- Síntese hidrotérmica de nanopartículas de In2O3 híbridos gêmeos heteroestruturas de ZnO de disco hexagonal para atividades fotocatalíticas aprimoradas e estabilidade