


MicroRNA-301 inibido restringe a angiogênese e o crescimento celular no carcinoma de células escamosas do esôfago por elevação do PTEN
Resumo
Objetivo
O carcinoma epidermóide de esôfago (CCEE) é caracterizado por metástase precoce e diagnóstico tardio. MicroRNA-301 (miR-301) é conhecido por participar de diversos tipos de câncer. No entanto, os efeitos do miR-301 no ESCC permanecem inexplorados. Assim, pretendemos explorar o papel do miR-301 na progressão do ESCC.
Métodos
A expressão de miR-301 e fosfatase e homólogo de tensina (PTEN) em tecidos ESCC e linhas celulares foi avaliada. Em seguida, as células selecionadas foram tratadas com miR-301 alterado ou oligonucleotídeo PTEN e plasmídeo e, em seguida, a capacidade de formação de colônias, viabilidade celular, migração, invasão, distribuição do ciclo celular e apoptose de células ESCC foram avaliadas. Além disso, o crescimento do tumor e a densidade dos microvasos (MVD) também foram avaliados, e a relação de direcionamento entre miR-301 e PTEN foi afirmada.
Resultados
MiR-301 foi regulado positivamente e PTEN foi regulado negativamente em tecidos e células ESCC. Células KYSE30 e células Eca109 foram selecionadas para ensaios funcionais. Em células KYSE30, o miR-301 inibido ou o PTEN superexpresso suprimiu os comportamentos malignos das células e o PTEN silenciado eliminou o impacto da inibição do miR-301 na progressão do ESCC. Em células Eca109, a superexpressão de miR-301 ou inibição de PTEN promoveu comportamentos malignos celulares, e a superexpressão de PTEN reverteu os efeitos da elevação de miR-301 na progressão de ESCC. O ensaio in vivo revelou que a inibição de miR-301 ou a superexpressão de PTEN reprimiu o crescimento do tumor ESCC e MVD, e a elevação de miR-301 ou redução de PTEN teve efeitos contrários. Além disso, PTEN foi alvo de miR-301.
Conclusão
Tomados em conjunto, os resultados do nosso estudo revelaram que o miR-301 afetou o crescimento celular, a metástase e a angiogênese por meio da regulação da expressão de PTEN em ESCC.
Introdução
O câncer de esôfago (CE), o 8º câncer mais comum em todo o mundo, é uma doença maligna crítica com alta mortalidade e prognóstico ruim [1]. Representando cerca de 90% do total de casos de CE, o carcinoma epidermóide de esôfago (CCEE) é a principal forma de CE na China [2]. Múltiplas causas, incluindo baixo nível socioeconômico, tabagismo, consumo de álcool, ingestão nutricional inadequada, alimentos ricos em nitrosaminas ou contaminados com micotoxinas, levam à ocorrência de ESCC [3]. Apesar dos resultados clínicos promovidos, bem como da administração, ainda há um prognóstico ruim entre os pacientes com CCEE, acompanhando uma taxa de sobrevida em 5 anos de 15–25% [4]. Portanto, é crucial confirmar oncogenes ou genes repressores de tumor que poderiam funcionar como biomarcadores no desenvolvimento de ESCC para fornecer métodos terapêuticos mais eficazes para pacientes com ESCC.
MicroRNAs (miRNAs) são pequenos RNAs não codificantes que desempenham um papel essencial na modulação da expressão gênica [5] e foram esclarecidos por terem a capacidade de influenciar a progressão do tumor por meio da regulação da estabilidade do mRNA e da capacidade dos mRNAs [6]. Uma quantidade de miRNAs como miR-4324 [7], miR-889-3p [8] e miR-9 [9] foram encontrados para estar associados ao processo de ESCC. MiR-301 é um membro dos miRNAs que é formado pela unidade de transcrição fam33a situada em 17q22-23 no genoma humano. A superexpressão de miR-301 foi identificada anteriormente, o que refletiu que está implicada em doenças humanas [6, 10]. No entanto, os mecanismos de função do miR-301 não foram descobertos no ESCC. Além disso, foi afirmado que o homólogo de fosfatase e tensina (PTEN) é freqüentemente interrompido em tumores e direcionado por mutações na linha germinativa em pacientes com câncer, o que desempenha um papel inibidor de tumores [11]. Foi validado que a desregulação de PTEN está correlacionada com o desenvolvimento de ESCC [12]. Curiosamente, uma pesquisa recente revelou que miR-301 tem como alvo PTEN no câncer de pulmão de células não pequenas [13]. No entanto, esta relação de segmentação entre miR-301 e PTEN no desenvolvimento de ESCC permanece a ser revelada. Nossa pesquisa se concentrou nos efeitos do miR-301 e do PTEN na progressão do ESCC, que permanecem em grande parte desconhecidos e são novidades. Inferimos que miR-301 pode influenciar a angiogênese e o crescimento celular em ESCC por meio da modulação da expressão de PTEN.
Materiais e métodos
Declaração de Ética
Consentimentos informados por escrito foram obtidos de todos os pacientes antes do estudo. Os protocolos deste estudo foram aprovados pelo Comitê de Ética do The Second Hospital of Jilin University e baseados nos princípios éticos para pesquisas médicas envolvendo seres humanos da Declaração de Helsinque. Os experimentos com animais foram estritamente consistentes com o Guia para o Manejo e Uso de Animais de Laboratório publicado pelo National Institutes of Health. O protocolo de experimentos com animais foi aprovado pelo Comitê de Uso e Cuidado Institucional de Animais do Segundo Hospital da Universidade de Jilin.
Assuntos de estudo
Cento e dez amostras de tecidos ESCC e tecidos normais adjacentes (> 5 cm do tumor) foram coletadas de pacientes ESCC (78 homens e 32 mulheres) que aceitaram esofagectomia no departamento de cirurgia torácica do Segundo Hospital da Universidade de Jilin em janeiro 2015 a dezembro de 2017. Entre os 110 pacientes, houve 84 casos> 60 anos e 26 casos ≤ 60 anos; tamanho do tumor:65 casos ≥ 5 cm e 45 casos <5 cm; 71 casos sem metástases em linfonodos (LNM) e 39 casos com LNM; o estágio de tumor, nódulo e metástase (TNM):60 casos estavam no estágio I + II e 50 casos no estágio III; localização do tumor:13 casos eram ESCC superior e 97 casos eram ESCC médio-inferior. Todos os pacientes foram diagnosticados com ESCC e não haviam aceitado radioterapia ou quimioterapia antes. Os tumores foram totalmente excisados e a margem cirúrgica negativa confirmada pelo anatomopatológico. De acordo com os critérios de estadiamento do ESCC propostos pela Union for International Cancer Control (UICC) em 2009 [14], o estadiamento patológico pós-operatório dos pacientes foi identificado como o estágio pT1-4N1-2 (I-IIIb). Não houve complicação significativa nos pacientes após a cirurgia, e os óbitos perioperatórios foram excluídos.
Reação em cadeia da polimerase quantitativa com transcrição reversa (RT-qPCR)
RNAs totais em tecidos e células foram extraídos usando kits Trizol (Invitrogen Inc., Carlsbad, CA, EUA). A concentração e a qualidade do RNA foram medidas. Os primers de RNA (Tabela 1) foram projetados e sintetizados por TaKaRa Biotechnology Co., Ltd. (Liaoning China). O RNA foi reversamente transcrito em cDNA com base nas instruções do kit de reagentes Takara PrimeScript ™ RT com g DNA Eraser (Takara). Realizamos qPCR no Light Cycler 480II (Roche) com o mix master Power PCR SYBR green PCR (Takara). U6 foi usado como o controle de carga de miR-301, e β-actina foi utilizada como a referência interna de PTEN. Os dados foram analisados usando 2 - △△ Ct método [15].
Análise de Western Blot
Tampão de lise RIPA (Beyotime Institute of Biotechnology, Shanghai, China) foi usado para extrair proteína total em células e tecidos, e a proteína foi quantificada por um kit BCA Protein Assay (Beyotime). A concentração de proteína de cada amostra foi medida e a eletroforese em gel de dodecilsulfato de sódio-poliacrilamida a 10% foi realizada. As amostras foram transferidas para as membranas de nitrocelulose, as quais foram então bloqueadas com leite em pó desnatado a 5% a 4 ° C durante a noite. Posteriormente, as membranas foram suplementadas com anticorpos primários PTEN e β-actina (ambos 1:500 e da Santa Cruz Biotechnology Inc, CA, EUA) para incubação durante a noite, em seguida, adicionados com os respectivos anticorpos secundários e incubados por 1 h. Depois de imersas no reagente quimioluminescente aprimorado (Pierce Chemical Inc., Dallas, TX, EUA) por 1 min, as membranas foram expostas em ambiente escuro e desenvolvidas usando o mini gerador de imagens de quimioluminescência LAS4000. Os valores de cinza foram avaliados por um software de sistema de imagem com β-actina como controle; portanto, foi obtida a proteína relativa final expressa. As bandas de proteína foram analisadas pelo software ImageJ2x.
Ensaio do gene Dual Luciferase Reporter
A sequência da região 3 'não traduzida (UTR) de PTEN foi prevista para interagir com miR-301, ou uma sequência mutada dentro dos locais alvo previstos foi sintetizada e inserida nos locais XbaI e FseI do vetor repórter de luciferase de controle pGL3 (Promega, WI, EUA). Em seguida, os vetores pGL3-PTEN-wt e pGL3-PTEN-mut foram produzidos. Os plasmídeos repórter de luciferase wt e mut identificados corretamente com miR-301 mímico e mímico NC foram co-transfectados em células KYSE30 e Eca109 por 48 h. Posteriormente, as células foram lisadas e as atividades da luciferase foram, respectivamente, determinadas por kits de detecção de luciferase (Promega).
Cultura celular, agrupamento e transfecção
As linhas celulares ESCC (KYSE-150, KYSE-30, Eca109 e KYSE-70) foram adquiridas no Instituto de Bioquímica e Biologia Celular de Xangai, Academia Chinesa de Ciências (Xangai, China) e as células epiteliais do esôfago humano (HEECs) foram adquiridas da Mingzhou Biotechnology Co., Ltd. (Zhejiang, China). As células foram cultivadas em meio RPMI 1640 (Invitrogen) suplementado com 10% de soro fetal bovino (FBS, Life Technologies, EUA), 100 unidades / ml de penicilina G de sódio (Sigma) e 100 μg / ml de sulfato de estreptomicina (Sigma). A expressão de MiR-301 e a expressão de mRNA de PTEN em cada linha celular foram medidas por RT-qPCR, e a linha celular com a maior e a menor expressão relativa foi escolhida para os experimentos celulares subsequentes.
As células KYSE-30 foram separadas em 7 grupos e, respectivamente, tratadas com inibidor miR-301, controle negativo do inibidor (NC), pcDNA-PTEN (denominado superexpresso (oe) -PTEN), pcDNA-NC (denominado oe-NC), inibidor miR-301 + pequeno RNA de interferência (si) -PTEN ou inibidor miR-301 + si-NC. As células Eca109 foram separadas em 7 grupos também e tratadas solidariamente com miR-301 mimetizador, mimetizador NC, si-PTEN, si-NC, miR-301 mimetizador + oe-PTEN, miR-301 mimetizador + oe-NC. inibidor NC, inibidor miR-301, miR-301 mimetizador, mimetizador NC, si-NC e si-PTEN foram adquiridos de GenePharma Ltd., Company (Shanghai, China); pcDNA-PTEN NC e pcDNA-PTEN foram obtidos de (Shanghai Sangon Bio-technology Corporation (Shanghai, China)). As células foram transfectadas transitoriamente em células ESCC por lipofectamina 2000 (Invitrogen) quando a confluência celular atingiu 60%.
Ensaio do kit de contagem de células (CCK-8)
As células foram semeadas em uma placa de 96 poços (1 × 10 3 células / poço) e incubados por diferentes períodos de tempo. Após incubado por 24 h, 48 h, 72 h e 96 h, cada poço foi suplementado com 10 μL de solução de CCK-8 (5 mg / mL) e, em seguida, as células de cada grupo foram incubadas a 37 ° C sem luz exposição por 2 h. Os valores de densidade óptica (DO) a 450 nm foram analisados por um leitor de microplacas (Bio-Rad Laboratories, Hercules, CA, EUA).
Ensaio de formação de colônia
As células foram semeadas a 500 células / poço em placas de 6 poços após a transfecção e cultivadas por 14 dias. As colônias foram fixadas com metanol, coradas com 0,5% de violeta de cristal e contadas ao microscópio invertido.
Ensaio Transwell
Células (5 × 10 3 ) incubados em meio RPMI 1640 foram semeados em câmaras apicais de dispositivos Transwell com membrana não revestida ou revestida com matrigel (Corning, NY, EUA). Após 24 h, as células nas câmaras apicais foram removidas, enquanto as células restantes na parte inferior foram fixadas e, em seguida, coradas com 0,1% de violeta de cristal. Um microscópio (Olympus Corporation, Tóquio, Japão) foi usado para contar em 3 campos aleatórios para calcular o número de células.
Citometria de fluxo
O ciclo celular e a apoptose foram avaliados por citometria de fluxo. Isotiocianato de fluoresceína de anexina V (10 µL) e iodeto de propídio (PI; 5 µL, Sigma) foram incubados com as células (5 × 10 5 células / poço) no escuro a 4 ° C por 30 min. A porcentagem de células apoptóticas foi calculada por meio de citômetro de fluxo (BD Biosciences, CA, EUA) com software FlowJo versão 10 (FlowJo LLC, OR, EUA).
Para avaliar o ciclo celular, as células (5 × 10 5 células / poço) foram fixados com etanol 75% durante a noite a 4 ° C e corados com 5 µl de PI / ribonuclease A (Sigma) a 4 ° C por 30 min no escuro. Os dados foram analisados com citômetro de fluxo (BD Biosciences). Sinais de fluorescência (14.000) de cada amostra foram coletados e calculados usando o software ModFit LT versão 3.2 (Verity Software House, Inc., ME, EUA).
Tumorigênese subcutânea em camundongos nus
Quarenta e dois camundongos nus BALB / c-nu fêmeas (4 anos de idade, pesando 16–24 g) foram obtidos do Experimental Animal Center Jilin University (Changchun, China). Os camundongos nus foram separados em 14 grupos ( n =3). Camundongos nus de sete grupos foram, respectivamente, injetados com células KYSE-30 de acordo com o agrupamento de células, e camundongos nus nos sete grupos restantes foram injetados separadamente com células Eca109 com base no agrupamento. A concentração de células KYSE-30 e Eca109 transfectadas foi ajustada para 5 × 10 6 células / 100 μL. Os camundongos nus foram fixados e injetados subcutaneamente com células ESCC correspondentes em condições estéreis. O maior comprimento (L) e largura (W) dos tumores foram medidos a cada semana e o volume do tumor ( V ) =1/2 × L × W 2 . Os ratos nus foram sacrificados no 5º th semana da injeção com os tumores ressecados, e os tumores foram pesados e fotografados. A taxa de formação de tumor foi calculada como o número de camundongos com tumor subcutâneo / número total de camundongos nus injetados no grupo × 100%. O tempo de injeção foi considerado em abcissa e o tamanho do tumor em ordenada; assim, a curva de crescimento do tumor foi representada graficamente.
Coloração imunohistoquímica
Os tecidos tumorais de camundongos nus foram fixados em formaldeído a 10%, embebidos em parafina e seccionados em 4 μm. Em seguida, as seções foram secas a 60 ° C por 2 h, desparafinadas por xileno, desidratadas por gradiente de etanol e incubadas com 50 μL 3% H 2 O 2 por 10 min. Posteriormente, as seções foram embebidas em solução tampão de ácido cítrico 0,01 M, fervida a 95 ° C por 20 min, bloqueada por solução de trabalho de soro de cabra normal a 37 ° C por 10 min e anexada com CD 34 (1:100, Santa Cruz) a 4 ° C durante a noite. Posteriormente, as seções foram suplementadas com polímero IgG de cabra anti-coelho / camundongo marcado com HRP (ZSGB-Bio, Pequim, China), contrastado por hematoxilina, desidratado e permeabilizado e então selado com bálsamo neutro. O PBS foi usado para substituir os anticorpos primários como o NC. Medição da densidade microvascular (MVD):os cortes foram observados em um microscópio de baixa ampliação. Uma célula endotelial ou um agrupamento de células endoteliais corado em amarelo acastanhado e significativamente distinguido com as células tumorais circundantes, e os tecidos conjuntivos foram considerados como um microvaso. A estrutura do ramo também foi considerada como um vaso se fosse desconectada, enquanto os vasos com tamanho do lúmen> 8 eritrócitos, camada muscular ou lúmen mais espesso foram excluídos. Os números de microvasos de 3 campos visuais altos foram registrados, e o número médio foi MVD de cada caso.
Análise estatística
Todas as análises estatísticas foram realizadas com o software SPSS versão SPSS 21.0 (IBM Corp. Armonk, NY, EUA) e apresentadas com Graphpad Prism Software 6.0. Os dados foram expressos como média ± desvio padrão. As diferenças entre dois grupos independentes foram testadas com o teste t de Student. ANOVA unilateral foi realizada para comparar três ou mais grupos. P valor <0,05 foi indicativo de diferença estatisticamente significativa.
Resultados
MiR-301 é altamente expresso, enquanto PTEN é mal expresso em tecidos e células ESCC
A expressão de MiR-301 e PTEN em tecidos ESCC e tecidos normais adjacentes foi avaliada usando RT-qPCR e análise de Western blot para revelar seus papéis em ESCC, e verificou-se que (Fig. 1a-c) miR-301 foi regulado positivamente, enquanto PTEN foi regulado para baixo em tecidos ESCC. Os pacientes foram divididos em grupos de baixa e alta expressão de acordo com o valor mediano de miR-301 ou expressão de PTEN para analisar a correlação entre a expressão de miR-301 ou PTEN e as características clínico-patológicas de pacientes com CCEE. Os resultados indicaram que a expressão de miR-301 / PTEN não foi relacionada com a idade, sexo, tamanho do tumor, localização e diferenciação, mas foi correlacionada com o estágio TNM e LNM de pacientes com CCEE (Tabela 2).
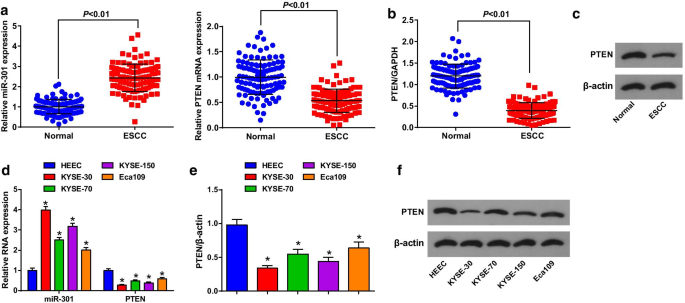
MiR-301 é altamente expresso, enquanto PTEN é mal expresso em tecidos e células ESCC. a Expressão de miR-301 e expressão de mRNA de PTEN no tecido ESCC detectado usando RT-qPCR; b expressão de proteína de PTEN no tecido ESCC detectada usando análise de Western blot; c bandas de proteína de PTEN no tecido ESCC em análise de Western blot; d expressão de miR-301 e expressão de mRNA de PTEN na linha de células ESCC detectada usando RT-qPCR; e expressão de proteína de PTEN na linha de células ESCC detectada usando análise de Western blot; f bandas de proteína de PTEN na análise de Western blot. * P <0,05 versus HEEC. Os dados foram expressos como média ± desvio padrão, e o teste t foi realizado para comparações entre dois grupos
Em seguida, a expressão de miR-301 e PTEN em 4 linhas de células ESCC e HEECs foi determinada usando RT-qPCR e análise de Western blot. Descobrimos que (Fig. 1d-f) miR-301 foi regulado positivamente e PTEN foi regulado negativamente em linhas de células ESCC, entre as quais KYSE-30 teve a expressão de miR-301 mais alta e a expressão de PTEN mais baixa, enquanto Eca109 teve a tendência contrária. Assim, a linha celular KYSE-30 foi tratada com miR-301 regulado negativamente / PTEN superexpresso e a linha celular Eca109 foi tratada com miR-301 superexpresso / PTEN silenciado nos experimentos celulares.
PTEN é direcionado por miR-301
Um software de bioinformática (http://www.microrna.org/) previu que PTEN era o gene alvo de miR-301 (Fig. 2a). Foi ainda confirmado por ensaio de gene repórter de luciferase dupla que a atividade da luciferase foi significativamente diminuída em células ESCC cotransfectadas com vetor PTEN-wt e miR-301 mimetizador em comparação com aqueles co-transfectados com vetor PTEN-mut e miR-301 mimetizador, implicando que miR-301 pode ligar-se particularmente a PTEN (Fig. 2b, c).
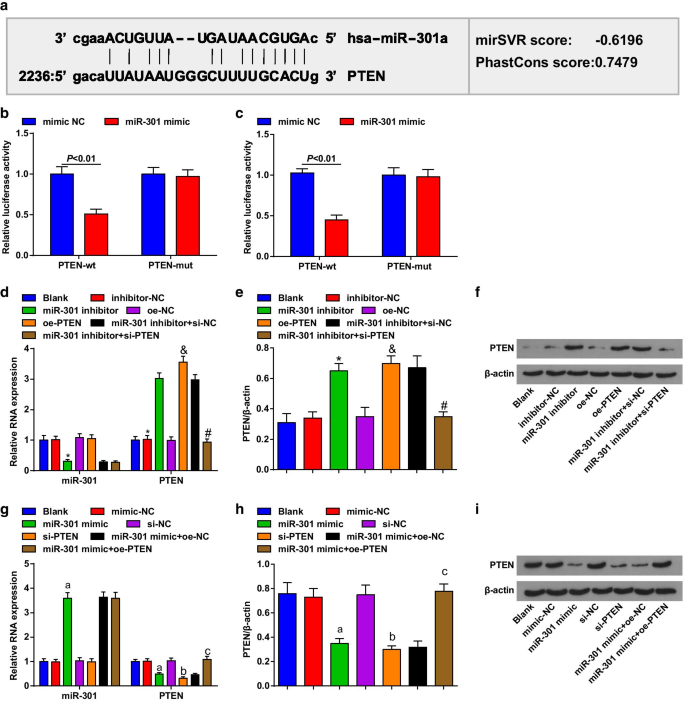
PTEN é o gene alvo de miR-301. a Os locais de ligação do miR-301 e PTEN foram previstos por software de previsão online; b a relação alvo entre miR-301 e PTEN em células KYSE-30 foi avaliada por ensaio de gene repórter de luciferase duplo; c a relação alvo entre miR-301 e PTEN em células Eca109 foi avaliada por ensaio de gene repórter de luciferase duplo; d expressão de miR-301 e expressão de mRNA de PTEN em células KYSE-30 detectadas usando RT-qPCR após miR-310 downregulation ou PTEN upregulation; e expressão de proteína de PTEN em células KYSE-30 detectada usando análise de Western blot após regulação negativa de miR-310 ou regulação positiva de PTEN; f bandas de proteína de PTEN em células KYSE-30 em análise de Western blot após miR-310 downregulation ou PTEN upregulation; g expressão de miR-301 e expressão de mRNA de PTEN em células Eca109 detectadas usando RT-qPCR após regulação positiva de miR-310 ou regulação negativa de PTEN; h expressão de proteína de PTEN em células Eca109 detectada usando análise de Western blot após regulação positiva de miR-310 ou regulação negativa de PTEN; eu bandas de proteína de PTEN em células Eca109 em análise de Western blot após regulação positiva de miR-310 ou regulação negativa de PTEN. * P <0,05 versus o grupo inibidor-NC, & P <0,05 versus o grupo oe-NC, # P <0,05 versus o inibidor miR-301 + grupo si-NC, um P <0,05 versus o grupo mímico-NC, b P <0,05 versus o grupo si-NC, c P <0,05 versus o grupo miR-301 mimetizador + oe-NC; N =3. Os dados foram expressos como média ± desvio padrão e o teste t foi realizado para comparações entre dois grupos. ANOVA foi usada para comparações entre vários grupos
RT-qPCR e análise de Western blot foram usados para avaliar a expressão de miR-301 e PTEN em células transfectadas, e verificou-se que em células KYSE-30 (Fig. 2d-f), células tratadas com inibidor de miR-301 desregulam miR-301 , enquanto PTEN regulado positivamente; as células tratadas com pcDNA-PTEN (oe-PTEN) aumentaram a expressão de PTEN e si-PTEN reverteram o efeito do inibidor miR-301 na expressão de PTEN. Em células Eca109 (Fig. 2g – i), as células tratadas com miR-301 mimetizam miR-301 com regulação positiva, enquanto PTEN com regulação negativa; células tratadas com si-PTEN diminuíram a expressão de PTEN, e pcDNA-PTEN (oe-PTEN) reverteu o papel inibidor de miR-301 mimetizador na expressão de PTEN. Esses dados sugeriram que o miR-301 tinha como alvo o PTEN.
miR-301 inibido ou PTEN superexpresso restringe a viabilidade de células ESCC; MiR-301 elevado ou PTEN reduzido promove a viabilidade de células ESCC
A viabilidade celular das células ESCC foi avaliada usando a formação de colônias e ensaios CCK-8. Os resultados revelaram que na linha celular KYSE-30 (Fig. 3a-c), a transfecção do inibidor miR-301 ou oe-PTEN reprimiu a capacidade de formação de colônias e a viabilidade celular; a transfecção de PTEN silenciado eliminou o impacto do inibidor miR-301 na viabilidade das células ESCC; na linha celular Eca109 (Fig. 3d-f), a transfecção de miR-301 mimetizador ou si-PTEN promoveu a capacidade de formação de colônias e a viabilidade celular; A superexpressão de PTEN reverteu o papel promotor da elevação de miR-301 na capacidade de formação de colônias e na viabilidade das células Eca109. Esses resultados sugeriram que o knockdown do miR-301 ou a superexpressão de PTEN reprimiu a viabilidade das células ESCC, que foram promovidas pela elevação do miR-301 ou inibição da PTEN.
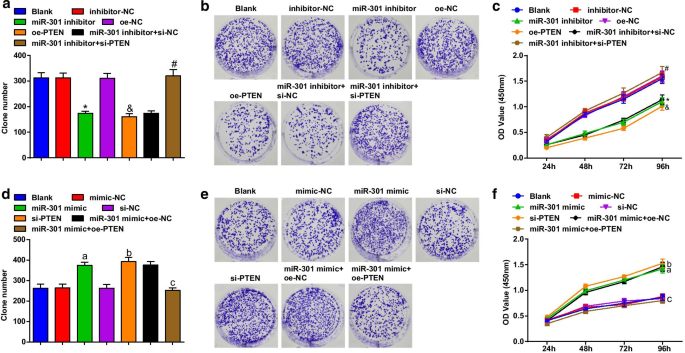
O miR-301 inibido ou PTEN superexpresso restringe a viabilidade das células ESCC; miR-301 elevado ou PTEN reduzido promove a viabilidade das células ESCC. a Número de colônias em células KYSE-30 após a transfecção detectada usando o ensaio de formação de colônia após a regulação negativa de miR-310 ou regulação positiva de PTEN; b capacidade de formação de colônia de células KYSE-30 após transfecção detectada usando ensaio de formação de colônia após regulação negativa de miR-310 ou regulação positiva de PTEN; c viabilidade de células KYSE-30 após transfecção detectada usando ensaio CCK-8 após regulação negativa de miR-310 ou regulação positiva de PTEN; d número de colônias em células Eca109 após a transfecção detectada usando ensaio de formação de colônias após regulação positiva de miR-310 ou regulação negativa de PTEN; e capacidade de formação de colônia de células Eca109 após transfecção detectada usando ensaio de formação de colônia após regulação positiva de miR-310 ou regulação negativa de PTEN; f viabilidade de células Eca109 após transfecção detectada usando ensaio CCK-8 após regulação positiva de miR-310 ou regulação negativa de PTEN; * P <0,05 versus o grupo inibidor-NC; & P <0,05 versus o grupo oe-NC; # P <0,05 versus o grupo inibidor miR-301 + si-NC; a P <0,05 versus o grupo NC-mimetizador; b P <0,05 versus o grupo si-NC; c P <0,05 versus o grupo miR-301 mimetizador + oe-NC, N =3. Os dados foram expressos como média ± desvio padrão, e ANOVA foi usada para comparações entre vários grupos
MiR-301 inibido ou PTEN superexpresso reprime a migração e invasão de células ESCC; MiR-301 elevado ou PTEN reduzido induz a migração e invasão de células ESCC
As habilidades de migração e invasão de células ESCC foram avaliadas usando o ensaio Transwell. Os resultados sugeriram que na linha celular KYSE-30 (Fig. 4a, b), a migração celular e as habilidades de invasão foram restringidas pela inibição de miR-301 ou superexpressão de PTEN; o papel supressor do inibidor miR-301 na migração celular e habilidades de invasão foi revertido por si-PTEN. Na linha celular Eca109 (Fig. 4c, d), a migração celular e as habilidades de invasão foram promovidas após a transfecção de miR-301 mimetizador ou si-PTEN; O PTEN superexpresso reverteu o impacto do miR-301 mimetizador na migração celular e nas habilidades de invasão. As descobertas acima implicaram que a migração e invasão de células ESCC foram inibidas pela repressão de miR-301 ou elevação de PTEN, enquanto foram facilitadas por regulação positiva de miR-301 ou regulação negativa de PTEN.
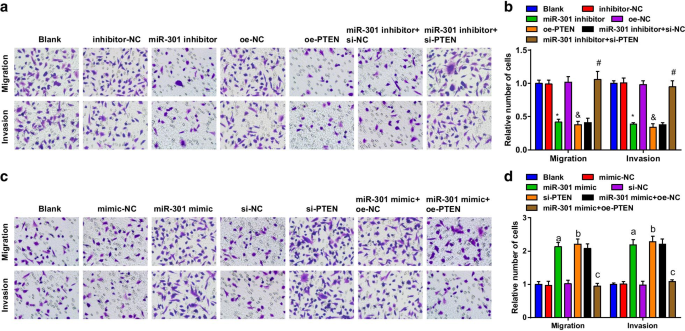
O miR-301 inibido ou PTEN superexpresso reprime a migração e invasão de células ESCC; miR-301 elevado ou PTEN reduzido promove migração e invasão de células ESCC. a Habilidades de migração e invasão de células KYSE-30 transfectadas avaliadas usando ensaio Transwell após regulação negativa de miR-310 ou regulação positiva de PTEN; b resultados estatísticos de migração e invasão em células KYSE-30 por meio de ensaio Transwell após regulação negativa de miR-310 ou regulação positiva de PTEN; c habilidades de migração e invasão de células Eca109 entre os grupos avaliados usando o ensaio Transwell após a regulação positiva de miR-310 ou regulação negativa de PTEN; d resultados estatísticos de migração e invasão em células Eca109 por meio da regulação positiva do ensaio Transwell miR-310 ou regulação negativa de PTEN. * P <0,05 versus o grupo inibidor-NC; & P <0,05 versus o grupo oe-NC; # P <0,05 versus o grupo inibidor miR-301 + si-NC; a P <0,05 versus o grupo NC-mimetizador; b P <0,05 versus o grupo si-NC; c P <0,05 versus o grupo miR-301 mimetizador + oe-NC, N =3. Os dados foram expressos como média ± desvio padrão, e ANOVA foi usada para comparações entre vários grupos
miR-301 inibido ou PTEN superexpresso induz parada do ciclo celular e apoptose de células ESCC; MiR-301 elevado ou PTEN reduzido suprime a parada do ciclo celular e apoptose de células ESCC
A citometria de fluxo foi usada para detectar a transição do ciclo celular e a apoptose das células após a transfecção, e os resultados indicaram que na linha celular KYSE-30 (Fig. 5a-d), a transfecção do inibidor miR-301 ou oe-PTEN promoveu a taxa apoptótica e aumento de células na fase G0 / G1, enquanto diminuiu nas fases S e G2 / M; a alteração da apoptose e parada do ciclo celular induzida pelo inibidor miR-301 poderia ser revertida por si-PTEN.
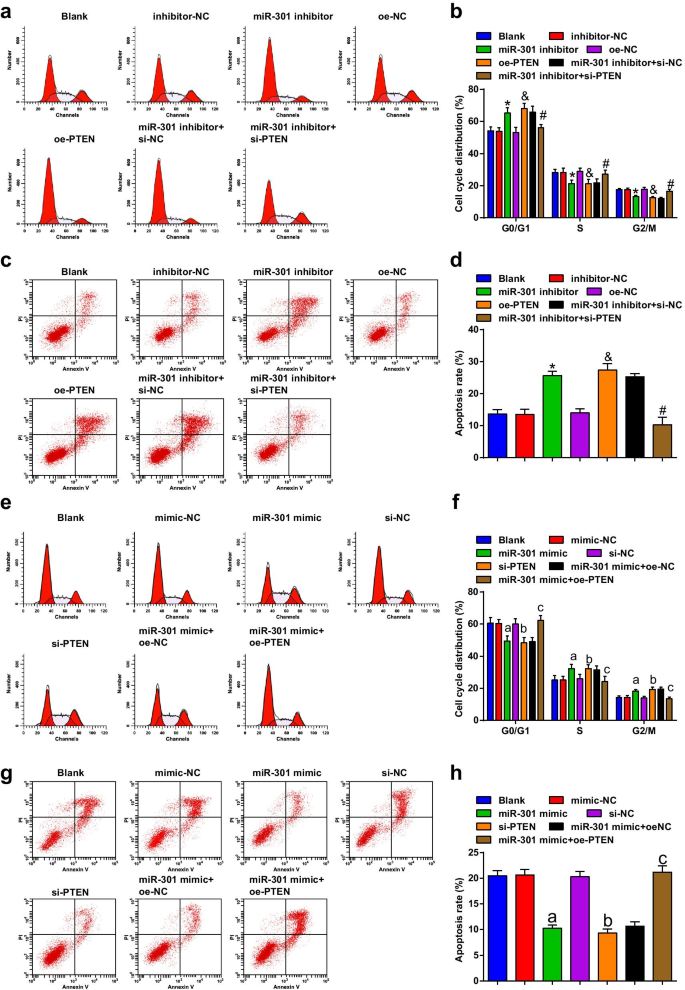
MiR-301 inibido ou PTEN superexpresso induz a parada do ciclo celular e apoptose de células ESCC; miR-301 elevado ou PTEN reduzido suprime a parada do ciclo celular e apoptose de células ESCC. a A distribuição do ciclo celular de células KYSE-30 em cada grupo foi detectada por citometria de fluxo após regulação negativa de miR-310 ou regulação positiva de PTEN; b resultados estatísticos de porcentagem nas fases G0 / G1, S e G2 / GM de células KYSE-30 em citometria de fluxo após regulação negativa de miR-310 ou regulação positiva de PTEN; c apoptose de células KYSE-30 foi detectada por citometria de fluxo após regulação negativa de miR-310 ou regulação positiva de PTEN; d taxa apoptótica de células KYSE-30 transfectadas detectadas usando citometria de fluxo após regulação negativa de miR-310 ou regulação positiva de PTEN; e a distribuição do ciclo celular de células Eca109 em cada grupo foi detectada por citometria de fluxo miR-310 regulação positiva ou regulação negativa de PTEN; f resultados estatísticos de porcentagem nas fases G0 / G1, S e G2 / GM de células Eca109 em citometria de fluxo após regulação positiva de miR-310 ou regulação negativa de PTEN; g apoptose de células Eca109 foi detectada por citometria de fluxo após regulação positiva de miR-310 ou regulação negativa de PTEN; h taxa apoptótica de células Eca109 transfectadas detectadas usando citometria de fluxo após regulação positiva de miR-310 ou regulação negativa de PTEN. * P <0,05 versus o grupo inibidor-NC; & P <0,05 versus o grupo oe-NC; # P <0,05 versus o grupo inibidor miR-301 + si-NC; a P <0,05 versus o grupo NC-mimetizador; b P <0,05 versus o grupo si-NC; c P <0,05 versus o grupo miR-301 mimetizador + oe-NC, N =3. Os dados foram expressos como média ± desvio padrão, e ANOVA foi usada para comparações entre vários grupos
De acordo com os resultados da citometria de fluxo, descobrimos que na linha celular Eca109 (Fig. 5e-h), a transfecção de miR-301 mimetizador ou si-PTEN inibiu a taxa apoptótica, diminuiu as células na fase G0 / G1 e aumentou a da Fase S e fases G2 / M; A superexpressão de PTEN reverteu o efeito do miR-301 mimetizador na taxa apoptótica e na parada do ciclo celular das células Eca109. Concluímos a partir desses resultados que miR-301 regulado negativamente ou PTEN regulado positivamente promoveu a transição do ciclo celular e apoptose em células ESCC, enquanto miR-301 inibido ou PTEN silenciado exerceu efeitos opostos.
MiR-301 inibido ou PTEN sobre-expresso restringe o crescimento do tumor e a angiogênese in vivo em ESCC; MiR-301 elevado ou PTEN reduzido aumenta o crescimento do tumor e angiogênese in vivo em ESCC
O crescimento e as mudanças de tumores ESCC em camundongos nus foram observados em cada grupo. O crescimento do tumor foi avaliado e os resultados implicaram que na linha celular KYSE-30 (Fig. 6a-e), camundongos nus injetados com inibidor miR-301 ou oe-PTEN reduziram o volume e o peso do tumor; o papel repressivo do inibidor miR-301 no crescimento do tumor foi abolido por si-PTEN. Na linha celular Eca109 (Fig. 6f-j), o volume do tumor e o peso do tumor foram avançados em camundongos nus injetados com miR-301 mimetizador ou si-PTEN; a superexpressão de PTEN reverteu o efeito de miR-301 mimetizador no crescimento do tumor. Enquanto isso, a expressão de CD34 em xenoenxertos de camundongos nus foi avaliada usando coloração imuno-histoquímica e os resultados mostraram que (Fig. 7a-d) em xenoenxertos KYSE-30, MVD foi contido após miR-301 downregulation ou PTEN upregulation; O PTEN silenciado reverteu o impacto da inibição do miR-301 no MVD. Em xenoenxertos Eca109, o MVD foi aumentado após a regulação positiva de miR-301 ou regulação negativa de PTEN; the enhancement of MVD induced by upregulated miR-301 could be abolished by overexpressed PTEN. These data indicated that miR-301 inhibition or PTEN overexpression repressed tumor growth and angiogenesis in ESCC, while miR-301 elevation or PTEN silencing had reverse effects.

Inhibited miR-301 or overexpressed PTEN restrains tumor growth in ESCC; elevated miR-301 or reduced PTEN increases tumor growth in ESCC. a Representative figures for the tumor growth observed by subcutaneous tumorigenesis in nude mice after KYSE-30 cells were transfected; b–d changes of tumor volume of each group after KYSE-30 cells were transfected; e changes of tumor weight of each group after KYSE-30 cells were transfected; f representative figures for the tumor growth observed by subcutaneous tumorigenesis in nude mice after Eca109 cells were transfected; g–i changes of tumor volume of each group after Eca109 cells were transfected; j changes of tumor weight of each group after Eca109 cells were transfected. * P < 0.05 versus the inhibitor-NC group; &P < 0.05 versus the oe-NC group; # P < 0.05 versus the miR-301 inhibitors + si-NC group; a P < 0.05 versus the mimic-NC group; b P < 0.05 versus the si-NC group; c P < 0.05 versus the miR-301 mimic + oe-NC group, n = 3 mice. Data were expressed as mean ± standard deviation, and ANOVA was used for comparisons among multiple groups

Inhibited miR-301 or overexpressed PTEN restrains angiogenesis in ESCC; elevated miR-301 or reduced PTEN increases angiogenesis in ESCC. a Representative images of tumor tissues observed by immunohistochemical staining in nude mice after KYSE-30 cells were transfected; b comparisons of MVD of KYSE-30 in tumor tissues among the groups; c representative images of tumor tissues observed by immunohistochemical staining in nude mice after Eca109 cells were transfected; d comparisons of MVD of Eca109 in tumor tissues among the groups *P < 0.05 versus the inhibitor-NC group; &P < 0.05 versus the oe-NC group; # P < 0.05 versus the miR-301 inhibitors + si-NC group; a P < 0.05 versus the mimic-NC group; b P < 0.05 versus the si-NC group; c P < 0.05 versus the miR-301 mimic + oe-NC group, n = 3 mice. Data were expressed as mean ± standard deviation, and ANOVA was used for comparisons among multiple groups
Discussion
EC is a kind of invasive malignancy in the gastrointestinal tract [16]. As the major type of EC, ESCC is a malignant tumor occurring in esophageal epithelial cells [17]. The miRNAs, known as small non-coding RNAs, have been demonstrated to function as a significant roles in leading molecules in the silencing of RNA [18]. Our research was designed to explore the effects of miR-301 and its target gene PTEN on ESCC progression, and we have found that the inhibited miR-301 could suppress angiogenesis and cell growth in ESCC by elevating PTEN.
MiR-301 expression was assessed, and we found that miR-301 was highly expressed in ESCC cells in comparison with HEEC, and the higher expression of miR-301 has also been found in ESCC tissues in contrast to the adjacent normal tissues. Similar to this result, Li et al. have identified that miR-301 presented high expression in myocardial infarction tissues [19]. In addition, we have elucidated that PTEN was targeted by miR-301, and the target relation has been pointed out by an extant literature [20]. We have also discovered that PTEN, which has been affirmed to be targeted by miR-301, was downregulated in both ESCC tissues and cells. Similarly, a previous research has unearthed that PTEN was poorly expressed in ESCC compared with non-tumor esophageal epithelial tissue [21]. Furthermore, Ma et al. have illuminated that PTEN expression was degraded in Eca109 cell line [22], which has also been selected for a series of experiments in this research. These studies provide evidence for the high expression of miR-301 and low expression of PTEN in ESCC.
Another important outcome in this research indicated that the inhibited miR-301 could repress the colony formation ability as well as the cell proliferation of ESCC cells via enhancing the PTEN expression, and elevated miR-301 or reduced PTEN had contrary effects. Similarly, Han et al. have elucidated that the downregulation of miR-301 mediated by luteolin has the ability to restrain the cell proliferation in prostate cancer [6]. A recent literature has revealed that the overexpression of PTEN suppresses the proliferation of pancreatic cancer cells [23], and a same result has been summarized in a study focusing on prostate cancer [24]. Besides, we have also unearthed that the downregulation of miR-301 or the elevation of PTEN could inhibit migration and invasion of ESCC cells, and elevated miR-301 or reduced PTEN exhibited the opposite trends. In accordance with this outcome, Shi et al. have supported that inhibited miR-301 attenuated migration and invasion of breast cancer cells [10], and it has been reported that the migration and invasion of ESCC cells could be repressed by the inhibition of miR-130b and the elevation of PTEN [25]. These publications helped verifying the oncogenic role of miR-301 and tumor-repressive effect of PTEN in diverse human cancers. Another result in our research was that inhibited miR-301 overexpressed PTEN to promote cell apoptosis and induce cell cycle arrest at the G0/G1 phase in ESCC cells, and elevated miR-301 or reduced PTEN had the inverse results. Similarly, it has been uncovered by a recent literature that activated PTEN induces cell cycle arrest and apoptosis in ESCC [26]. Consistently, Tian et al. have found in their study that the elevation of PTEN inhibited the angiogenesis by reducing the expression of vascular endothelial growth factor in hepatocellular carcinoma [27]. Based on the above data, the roles of miR-301 and PTEN in cell apoptosis and angiogenesis in diverse diseases were further confirmed. Consequently, we concluded that the downregulation of miR-301 could restrain the tumor growth in ESCC through the high expression of PTEN, and the similar conclusion has also been unveiled in breast cancer [10] and prostate cancer [28]. On the contrary, miR-301 elevation or PTEN reduction induced the tumor growth in ESCC. It could be concluded that miR-301 and PTEN participated in the in vivo cancer cell growth.
Conclusion
In this study, we have shown that the repression of miR-301 prohibits angiogenesis, cell proliferation, migration and invasion but promotes apoptosis in ESCC cells by upregulating PTEN. This research may further the understanding on potential molecular mechanisms of ESCC and provide novel targets for ESCC treatment.
Abreviações
- ESCC:
-
Esophageal squamous cell carcinoma
- PTEN:
-
Phosphatase and tensin homologue
- MVD:
-
Microvessel density
- EC:
-
Esophageal cancer
- miRNAs:
-
MicroRNAs
- LNM:
-
Lymph node metastasis
- UICC:
-
Union for International Cancer Control
- RT-qPCR:
-
Reverse transcription quantitative polymerase chain reaction
- 3′UTR:
-
3′-Untranslated region
- WT:
-
Wild type
- MUT:
-
Mutant type
- HRP:
-
Peroxidase de rábano
- FBS:
-
Soro fetal bovino
- OE:
-
Overexpressed
- NC:
-
Negative control
- CCK-8:
-
Cell counting kit
- DMEM:
-
Meio Eagle modificado de Dulbecco
- PI:
-
Propidium iodide
- FITC:
-
Fluorescein isothiocyanate
- MVD:
-
Microvessel density
- ANOVA:
-
Analysis of variance
Morfologia de superfície regulada de nanofibras compostas de polianilina / ácido polilático por meio de vários dopagem de ácidos inorgânicos para aumentar a biocompatibilidade em engenharia de t…
SOX2 regula o eixo lncRNA CCAT1 / MicroRNA-185-3p / FOXP3 para afetar a proliferação e auto-renovação de células-tronco de câncer cervical
Nanomateriais
- A regulação negativa de microRNA-342-5p ou a regulação positiva de Wnt3a inibe a angiogênese e mantém a estabilidade da placa aterosclerótica em camundongos com aterosclerose
- Preparação de nanopartículas de mPEG-ICA carregadas com ICA e sua aplicação no tratamento de dano celular H9c2 induzido por LPS
- Análise de Actina e Organização de Adesão Focal em Células U2OS em Nanoestruturas Poliméricas
- As Nanofibras Hierárquicas de Poliamida 6-ZnO Antibacteriana Fabricadas por Deposição de Camada Atômica e Crescimento Hidrotérmico
- Estudo in vitro da influência das nanopartículas de Au nas linhas celulares HT29 e SPEV
- Promoção do crescimento celular SH-SY5Y por nanopartículas de ouro modificadas com 6-mercaptopurina e um peptídeo penetrador de neurônio
- Nanopartículas de maghemita atuam como nanozimas, melhorando o crescimento e a tolerância ao estresse abiótico em Brassica napus
- Nanopartículas de ouro de formato redondo:efeito do tamanho da partícula e da concentração no crescimento da raiz de Arabidopsis thaliana
- Novo suporte de catalisador anódico para célula de combustível de metanol direto:caracterizações e desempenho de célula única
- 5G e o desafio do crescimento exponencial de dados