


SOX2 regula o eixo lncRNA CCAT1 / MicroRNA-185-3p / FOXP3 para afetar a proliferação e auto-renovação de células-tronco de câncer cervical
Resumo
Foi apresentado o papel dos longos RNAs não codificantes (lncRNAs) no câncer cervical (CC). Nosso objetivo é discutir o efeito da região determinante do sexo Y-box 2 (SOX2) / lncRNA do transcrito associado ao câncer de cólon (CCAT1) / microRNA-185-3p (miR-185-3p) / proteína da caixa forkhead 3 (FOXP3 ) sobre a capacidade de proliferação e auto-renovação das células-tronco CC. As expressões de MiR-185-3p, SOX2, CCAT1 e FOXP3 foram testadas em tecidos e células CC. A relação entre a expressão de SOX2 / CCAT1 e características clínico-patológicas em pacientes com CC foi verificada. As investigações de perda e ganho de função foram conduzidas em CD44 + Células HeLa para discutir funções biológicas e capacidade de auto-renovação. Finalmente, as relações entre SOX2, CCAT1, FOXP3 e miR-185-3p foram verificadas. A expressão de miR-185-3p foi diminuída, enquanto as expressões de SOX2, CCAT1 e FOXP3 aumentaram em tecidos e células de CC. As expressões de SOX2 e CCAT1 foram associadas ao tamanho do tumor, metástases em linfonodos e a federação internacional de ginecologia e estágio obstétrico de CC. A regulação negativa de SOX2 ou CCAT1 e a regulação positiva de miR-185-3p resultaram na inibição da proliferação, invasão, migração e número de esfera celular, bem como aceleração de apoptose de CD44 + Células HeLa. SOX2 poderia se ligar a CCAT1 que afetou a expressão de miR-185-3p, e FOXP3 foi direcionado por miR-185-3p.
Introdução
O câncer cervical (CC) é a quarta maior causa de mortalidade em mulheres, com uma estimativa de 570.000 casos e 311.000 mortes em todo o mundo em 2018 [1]. Esta doença complexa participa de uma variedade de fatores, incluindo efeitos genéticos e infecção viral [2]. Com o desenvolvimento do co-teste do papilomavírus humano e da vacinação do papilomavírus humano, os procedimentos diagnósticos precoces da displasia cervical e do câncer resultam em uma redução na incidência, morbidade e mortalidade do CC [3]. Para pacientes com CC inicial, a cirurgia é recomendada, como cirurgia de preservação de fertilidade, biópsia em cone, traquelectomia radical, dissecção de linfonodo pélvico, radioterapia pélvica e braquiterapia [4]. Devido à metástase ou recorrência em pacientes com CC avançado, o prognóstico ainda é ruim [5]. Portanto, ainda é urgente identificar novos e eficazes marcadores prognósticos e estratégias terapêuticas para melhorar o tratamento do CCU.
A região determinante do sexo Y-box 2 (SOX2) é um membro crítico da família do fator de transcrição SOX e se manifesta principalmente em células-tronco embrionárias e adultas e também expresso em células-tronco tumorais [6]. Ele revelou que o SOX2 modula a radiorresistência em CC pela via de sinalização hedgehog [7]. Outro estudo demonstrou que SOX2 é crucial para manter a subpopulação de células-tronco cancerosas em linhagens de células CC [8]. RNAs não codificantes longos (lncRNAs) são uma classe de moléculas de RNA com 200 nucleotídeos de comprimento [9]. O transcrito-1 associado ao câncer de cólon LncRNA (CCAT1) está localizado no cromossomo humano 8q24.21 e é considerado um "ponto quente" que resulta em mutações genéticas no câncer de cólon [10]. Um estudo relatou que o CCAT1 acelera a proliferação celular e a invasão do CC [11]. De acordo com Jia et al., CCAT1 aumenta dramaticamente a proliferação, migração e invasão de células CC [12]. Além disso, outro estudo revelou que o CCAT1 aumenta o grau de malignidade da doença inflamatória intestinal por meio da destruição da barreira intestinal, reduzindo o microRNA-185-3p (miR-185-3p) [13]. MiRNA pode controlar reversamente a expressão do gene através da redução do mRNA e suprimindo a tradução [14]. Experimentos in vitro em um estudo anterior revelaram que miR-185-3p modula a radiorresistência do carcinoma nasofaríngeo [15]. Outro estudo implicou que o miR-185 participa da resistência à cisplatina do câncer de ovário in vivo e in vitro [16]. A proteína 3 da caixa forkhead (FOXP3) é um fator de transcrição pertencente à família de proteínas FOX, que é encontrada pela primeira vez em células T reguladoras (Treg) e desempenha um papel vital na manutenção e no processo das células Treg [17]. Um estudo relata que o FOXP3 está conectado à linfangiogênese do CC [18]. Outro estudo revela que o nível de FOXP3 está dramaticamente ligado à federação internacional de estágio de ginecologia e obstetrícia (estágio de FIGO) e tamanho do tumor de CC [19]. Neste estudo, examinamos os efeitos do eixo SOX2 / CCAT1 / miR-185-3p / FOXP3 na capacidade de proliferação e autorrenovação de células-tronco CC.
Materiais e métodos
Aprovação Ética e Consentimento para Participar
Os experimentos envolvendo seres humanos foram realizados em conformidade com os princípios expressos na Declaração de Helsinque. O estudo foi aprovado pelo Comitê de Ética em Pesquisa do The First Hospital of Jilin University. Todos os participantes assinaram um termo de consentimento livre e esclarecido.
Assuntos de estudo
De dezembro de 2016 a dezembro de 2018, 39 casos de tecidos CC e tecidos normais adjacentes correspondentes foram colhidos de pacientes com CC e preservados em nitrogênio líquido. Os critérios de inclusão foram os seguintes:(1) As pacientes foram confirmadas como CC por patologia de biópsias em canais transcervicais e cervicais, citologia de esfregaço cervical, teste de iodo cervical, espéculo vaginal e ressecção de vértebra cervical. (2) Os pacientes não receberam radioterapia e quimioterapia 2 semanas antes da operação. Os critérios de exclusão foram os seguintes:(1) pacientes submetidos a qualquer tipo de radioterapia ou quimioterapia, (2) pacientes discordantes da coleta da amostra e (3) pacientes com distúrbios do sistema imunológico.
Seleção e cultura de células
As linhas celulares CC (SiHa, HeLa, CaSki, HCC94 e C33A) e a linha celular epitelial cervical humana imortalizada H8 foram adquiridas em Shanghai Bioleaf Biotech Co., Ltd. (Shanghai, China). Células CC SiHa, HeLa e HCC94 foram cultivadas em meio de Eagle modificado por Dulbecco de alta glicose (DMEM) contendo 10% de soro fetal bovino (FBS), enquanto as células CaSki, C33A e H8 em meio Roswell Park Memorial Institute (RPMI) -1640 contendo 10 % FBS (37 ℃ e 5% CO 2 ) As células foram separadas e subcultivadas a cada 2 dias.
Classificação de células-tronco CC (CD44 + Células HeLa)
As células-tronco CC foram separadas da linha de células CC HeLa por cultura em suspensão de células. As células CC HeLa foram cultivadas por meio de Dulbecco modificado Iscove isento de soro (IMDM) em uma placa de Petri de adesão ultrabaixa por 21 d com o meio semiquantitativamente trocado a cada 3-5 d. Algumas células foram suspensas em esferas, e células formadoras de esferas HeLa (SFCs) foram obtidas. A propriedade das células esféricas foi identificada e analisada. As células foram separadas por tripsina e ajustadas para 1 × 10 6 células / mL. As células foram adicionadas com anticorpo CD44 e classificadas por citometria de fluxo. As células HeLa com CD44 positivo eram células-tronco tumorais HeLa, enquanto com CD44 negativo eram células não-tronco HeLa. As células-tronco CC foram cultivadas em DMEM / F12 e anexadas com 20 ng / mL de fator de crescimento de fibroblastos básico (bFGF), 20 ng / mL de fator de crescimento epidérmico (EGF) e B27. O meio continha 1% de penicilina e estreptomicina [20].
Tratamento celular
CD44 + As células HeLa foram transfectadas com sh-SOX2, sh-SOX2 controle negativo (NC), sh-CCAT1, sh-CCAT1 NC, miR-185-3p mimetizador, mimetizador NC, sh-CCAT1 e inibidor miR-185-3p, bem como sh-CCAT1 e inibidor NC. Todas as sequências de oligonucleotídeos foram fornecidas pela GenePharma (Shanghai, China). Separadas pela tripsina, as células foram semeadas em uma placa de 6 poços com 3 × 10 6 células / poço. Quando atingiu 60% de confluência, as células foram alteradas para meio sem soro e incubadas por 1 h. A transfecção foi facilitada pelo reagente de transfecção Lipofectamine 2000 (Invitrogen, Carlsbad, Califórnia, EUA).
Reação em cadeia da polimerase quantitativa com transcrição reversa (RT-qPCR)
O RNA em tecidos e células foi extraído por Trizol (Invitrogen). O RNA (1 μg) foi revertido em cDNA pelo kit RTase do vírus da leucemia murina Moloney (Invitrogen). O cDNA foi adicionado ao sistema de PCR em tempo real. Os primers foram desenvolvidos por Shanghai Sangon Biotechnology Co., Ltd. (Shanghai, China) (Tabela 1). U6 foi o controle de carga de miR-185-3p, enquanto gliceraldeído fosfato desidrogenase (GAPDH) de SOX2, CCAT1 e FOXP3. Os resultados foram analisados por 2 −ΔΔCt método.
Ensaio de Western Blot
A proteína total nas células e tecidos foi extraída. A concentração de proteína foi determinada por kits de ácido bicinconínico (AmyJet Scientific, Wuhan, Hubei, China). A proteína foi misturada com tampão de carga e fervida por 5 min, seguida por banho de gelo e centrifugação. A proteína foi tratada com eletroforese em gel de poliacrilamida dodecil sulfato de sódio a 10% e transferida para uma membrana. A membrana foi bloqueada com 5% de leite desnatado por 1 h, sondado com anticorpos primários SOX2 (1:1000, Jiangsu Rui sitan Co., Ltd., Jiangsu, China), FOXP3 (1:1000, Abcam Inc., Cambridge, MA , EUA), GAPDH (1:1000, Cell Signaling Technology, Beverly, MA, EUA) e re-sondado com anticorpo secundário marcado por peroxidase de rábano, coberto por filme conservante e exposto. O imager Bio-rad Gel Doc EZ (Bio-rad, Califórnia, EUA) foi adotado para o desenvolvimento. As imagens das proteínas foram analisadas pelo software ImageJ2x.
Ensaio de kit de contagem de células (CCK) -8
O ensaio CCK-8 foi implementado com os kits (Beyotime, Xangai, China). Células (1 × 10 4 ) foram semeados em uma placa de 96 poços e incubados. Cultivadas por 0, 24, 48 e 72 h, as células foram adicionadas com 10 μL / poço de solução de CCK-8 e incubadas por 1 h. O valor de densidade óptica foi determinado com leitor de microplaca de comprimento de onda completo Multiskan Spectrum a 450 nm. Seis poços foram tomados para contar o valor médio. A curva de crescimento celular foi traçada com o tempo como ordenada e a viabilidade celular relativa como ordenada. O valor da densidade óptica representou a proliferação celular.
Citometria de fluxo
Células (1 × 10 6 ) foram centrifugados a 1500 r / min, suspensos com 200 μL de tampão de ligação, incubados com 5 μL de iodeto de propídio (PI) e 5 μL de isotiocianato de fluoresceína V-Anexina (FITC) por sua vez e anexados com 400 μL de tampão. A taxa de apoptose celular foi verificada por citômetro de fluxo (BD Biosciences, NJ, EUA).
Teste Scratch
As células transfectadas foram separadas e preparadas em suspensão de células. Suspensões de células (1 × 10 6 células / mL) foram semeados em uma placa de 6 poços e cultivados até 80-90% de confluência. Arranhões transversais e longitudinais na placa de 6 poços foram feitos uniformemente por uma ponta de pipeta asséptica de 10 μL ao longo da régua. O meio original foi substituído por meio completo e as células foram cultivadas continuamente. A migração das células foi observada ao microscópio às 0 he 72 he fotografada no mesmo local.
Ensaio Transwell
As células transfectadas foram tripsinizadas, centrifugadas a 1000 ge enxaguadas com DMEM. Uma câmara Transwell totalmente hidratada foi colocada em uma placa de 24 poços contendo 10% de FBS-DMEM (600 mL / poço). Suspensão de células (5 × 10 5 células / mL, 200 μL) foi anexado na câmara superior Transwell (revestido com 1:8 Matrigel 80 μL), enquanto 500 μL 20% FBS-DMEM para a câmara inferior. As células foram cultivadas continuamente por 24 h, fixadas com 500 μL de paraformaldeído a 4% (PFA) e coradas com solução de coloração com violeta de cristal a 0,1%. Em seguida, as células na superfície e na borda da câmara superior foram eliminadas com um cotonete. Cinco campos foram selecionados aleatoriamente e as células foram contadas em um microscópio Nikon Eclipse TE2000-S (Nikon, Japão).
Ensaio de formação de esfera
As células foram semeadas em uma placa de 6 poços de baixa adsorção contendo meio de suspensão isento de soro com 200 células / poço. Após 2 w, a taxa de formação de esferas de células foi observada sob um microscópio Nikon Eclipse TE2000-S (Nikon), e a taxa de formação de esferas foi calculada como o número médio de esferas / número de células semeadas × 100%.
Ensaio de imunoprecipitação da cromatina (ChIP)
O ensaio ChIP foi operado com o kit ChIP (Upstate, NY, EUA). Os anticorpos SOX2 (1:1000, Re-stem Biotech, Jiangsu, China) e IgG de coelho normal (12–370, Millipore, EUA) foram utilizados para imunoprecipitar o complexo reticulado proteína-DNA. CD44 + As células HeLa foram fixadas com PFA a 1% e incubadas para produzir reticulação DNA-proteína. Em seguida, o DNA foi cortado em fragmentos de cromatina de 200-300 bp por ultrassom. O DNA da cromatina precipitado foi recuperado e analisado por RT-qPCR.
Ensaio pull-down de RNA
Os plasmídeos de tipo selvagem (WT) e tipo mutante (MUT) marcados com biotina (50 nM) foram transfectados em CD44 + Células HeLa, respectivamente. As células foram incubadas com lisado celular específico (Ambion, Austin, Texas, EUA) após 48 h de transfecção. O lisado celular (50 mL) foi subempacotado. O lisado residual foi eclodido com esferas de estreptavidina M-280 (Sigma, St. Louis, MO, EUA) pré-revestidas com ARNt isento de RNase e de levedura (Sigma). Em seguida, as células foram limpas duas vezes com lisado frio, três vezes com tampão de baixo teor de sal e uma vez com tampão de alto teor de sal. Uma sonda miR-185-3p antagonista foi configurada como NC. O RNA total foi extraído por Trizol, e o nível de CCAT1 foi testado por RT-qPCR.
Ensaio do gene Dual Luciferase Reporter
Os locais de ligação potenciais E1 e E2 de SOX2 na região do promotor CCAT1 foram previstos por https://jaspar.genereg.net/. A sequência do promotor CCAT1 contendo SOX2 e o sítio de ligação E1 de CCAT1 foi sintetizada, e os vetores CCAT1 3′UTR WT (E1-WT) e CCAT1 3′UTR MUT (E1-MUT) foram formados. Os vetores foram clonados em pmirGLO (Beyotime). Depois disso, CCAT1-WT / pmirGLO ou CCAT1-MUT / pmirGLO foi co-transfectado com sh-SOX2 ou sh-SOX2 NC para CD44 + Células HeLa, respectivamente, por 2 dias e depois lisadas. A atividade da luciferase foi testada pelo sistema de detecção de luciferase (Takara, Dalian, China).
O site de bioinformática foi utilizado para prever e analisar os sítios de ligação de CCAT1 e miR-185-3p. Os locais de ligação de CCAT1 e miR-185-3p foram verificados por ensaio de gene repórter de luciferase duplo. Foi composto CCAT1 3′UTR contendo o sítio de ligação miR-185-3p. CCAT1 3′UTR WT e CCAT1 3′UTR MUT foram construídos e co-transfectados com mímicos NC e mímicos miR-185-3p para CD44 + Células HeLa por 2 d. Em seguida, as células foram lisadas e a atividade da luciferase foi testada pelo sistema de detecção de luciferase (Takara). O mesmo método foi aplicado para verificar a relação de direcionamento entre miR-185-3p e FOXP3.
Análise estatística
Todos os dados foram avaliados pelo software SPSS 21.0 (IBM Corp. Armonk, NY, EUA). Os dados de medição foram indicados como média ± desvio padrão. O teste t foi aplicado para discrepância entre dois grupos e análise de variância unilateral (ANOVA) seguido pelo teste de comparações múltiplas de Tukey para discrepância entre os grupos. A variável de classificação foi avaliada pelo teste exato de Fisher. A p valor inferior a 0,05 foi considerado significativo.
Resultados
miR-185-3p A expressão diminui, enquanto as expressões SOX2, CCAT1 e FOXP3 aumentam nos tecidos CC e as expressões SOX2 e CCAT1 estão vinculadas ao tamanho do tumor, linfonodo Metástase (LNM) e estágio FIGO
Ao detectar o papel do eixo SOX2 / CCAT1 / miR-185-3p / FOXP3 na capacidade de proliferação e autorrenovação de células-tronco CC, as expressões de miR-185-3p, SOX2, CCAT1 e FOXP3 em tecidos CC e tecidos normais adjacentes foram testado por RT-qPCR e ensaio Western blot. Foi manifestado que (Fig. 1a – c) a expressão de miR-185-3p foi reduzida, enquanto as expressões de SOX2, CCAT1 e FOXP3 foram aumentadas em tecidos CC.
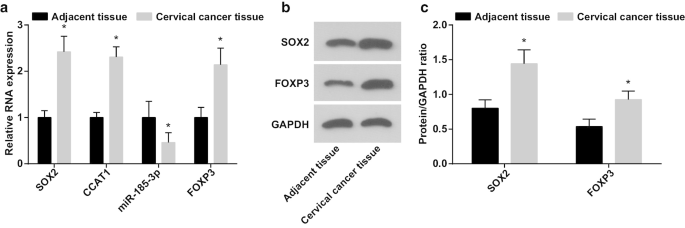
A expressão de miR-185-3p diminui, enquanto as expressões de SOX2, CCAT1 e FOXP3 aumentam em tecidos CC. a Comparação da expressão de mRNA de SOX2, CCAT1, miR-185-3p e FOXP3 mRNA em CC e tecidos normais adjacentes. b Bandas de proteínas da expressão das proteínas SOX2 e FOXP3 em CC e tecidos normais adjacentes. c Comparação da expressão das proteínas SOX2 e FOXP3 em CC e tecidos normais adjacentes. * p <0,05 vs. tecidos normais adjacentes. Os dados de medição foram representados como média ± desvio padrão, e as comparações entre dois grupos foram avaliadas pelo teste t
A relação entre a expressão de SOX2 / CCAT1 e características clínico-patológicas do CC foi analisada (Tabela 2). SOX2 e CCAT1 superexpressos em CC foram conectados com o tamanho do tumor, LNM e estágio FIGO, indicando que as expressões de SOX2 e CCAT1 foram maiores em pacientes com tamanho de tumor maior, LNM e estágio avançado de FIGO de pacientes com CC.
miR-185-3p a expressão reduz, enquanto as expressões SOX2, CCAT1 e FOXP3 aumentam em CD44 + Células HeLa
Em seguida, as expressões miR-185-3p, SOX2, CCAT1 e FOXP3 em células epiteliais cervicais humanas imortalizadas H8 e linhas de células CC SiHa, CasKi, HeLa, HCC94 e C33A foram testadas. Foi sugerido que (Fig. 2a – c) a expressão de miR-185-3p foi degradada, enquanto SOX2, CCAT1 e FOXP3 foram elevados em linhas de células CC. Entre eles, SOX2 / CCAT1 / miR-185-3p / FOXP3 em células HeLa tiveram a maior diferença de expressão em células H8; assim, as células HeLa foram selecionadas e classificadas; HeLa-SFCs foram obtidos por culturas em suspensão de células, e a expressão de CD44 na superfície celular antes e após a classificação foi identificada por citometria de fluxo. Os resultados destacaram que a taxa positiva de CD44 em HeLa-SFCs foi marcadamente maior do que antes da classificação, sugerindo uma classificação bem-sucedida de células-tronco CC (Fig. 2d). As células-tronco classificadas foram nomeadas como CD44 + Células HeLa. Em seguida, as expressões miR-185-3p, SOX2, CCAT1 e FOXP3 foram testadas em HeLa e CD44 + Células HeLa (Fig. 2e – g). Foi manifestado que as expressões SOX2, CCAT1 e FOXP3 foram reguladas para cima e miR-185-3p foi regulado para baixo em CD44 + Células HeLa.
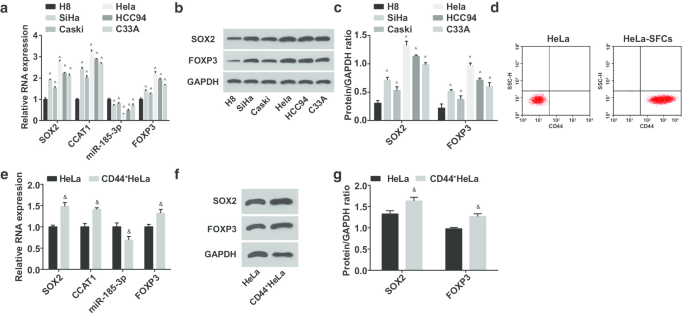
A expressão miR-185-3p reduz, enquanto as expressões SOX2, CCAT1 e FOXP3 aumentam em CD44 + Células HeLa. a Comparação da expressão de mRNA de SOX2, CCAT1, miR-185-3p e FOXP3 em linhas de células H8 e CC. b Bandas de proteínas da expressão das proteínas SOX2 e FOXP3 em linhas de células H8 e CC. c Comparação da expressão das proteínas SOX2 e FOXP3 em linhas de células H8 e CC. d Detecção da taxa de expressão de CD44 em células HeLa antes e depois da classificação por citometria de fluxo. e Comparação da expressão de mRNA SOX2, CCAT1 e miR-185-3p entre células HeLa e CD44 + Células HeLa. f Banda de proteína da expressão da proteína SOX2 em células HeLa e CD44 + Células HeLa. g Comparação da expressão da proteína SOX2 entre células HeLa e CD44 + Células HeLa. ^ p <0,05 vs. células H8. e p <0,05 vs. células HeLa. Os dados de medição foram descritos como média ± desvio padrão, as comparações entre dois grupos foram avaliadas pelo teste t e as comparações entre vários grupos foram avaliadas por ANOVA de uma via seguida pelo teste post hoc de Tukey
Regulagem descendente de SOX2 e regulagem negativa de CCAT1 Diminui a proliferação, migração, invasão e taxa de formação de esfera e aumenta a apoptose de CD44 + Células HeLa
Posteriormente, SOX2 e CCAT1 foram silenciados em CD44 + Células HeLa para explorar seus efeitos na proliferação e autorrenovação de células-tronco CC. Detectado por ensaio CCK-8, citometria de fluxo, teste de raspagem, ensaio Transwell e experimento de formação de esfera, a proliferação, migração, invasão e taxa de formação de esfera foram suprimidas e a apoptose de CD44 + As células HeLa foram promovidas pela inibição de SOX2 e CCAT1 (Fig. 3a – i). Foi indicativo que o silenciamento de SOX2 ou CCAT1 inibiu a proliferação e autorrenovação de células-tronco CC.
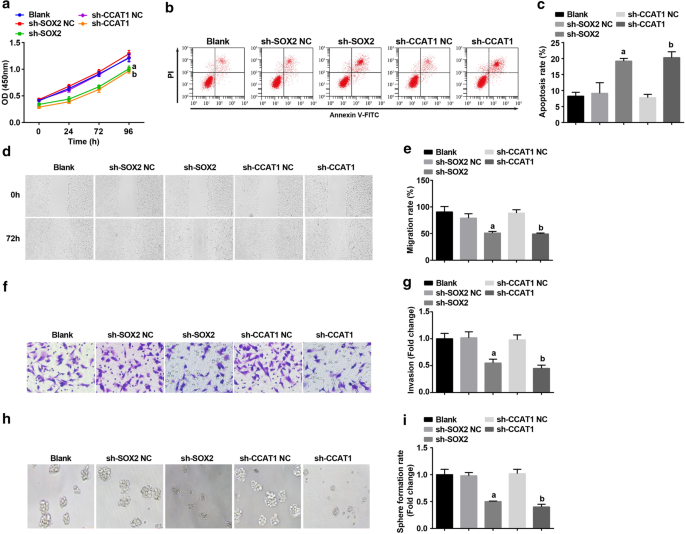
A regulação negativa de SOX2 e a regulação negativa de CCAT1 diminuem a proliferação, migração, invasão e taxa de formação de esferas e aumentam a apoptose de CD44 + Células HeLa. a O ensaio CCK-8 testou a curva de crescimento celular em células tratadas com sh-CCAT1 ou sh-SOX2. b A citometria de fluxo detectou apoptose celular em células tratadas com sh-CCAT1 ou sh-SOX2. c Comparação da taxa de apoptose celular em células tratadas com sh-CCAT1 ou sh-SOX2. d Migração de células em células tratadas com sh-CCAT1 ou sh-SOX2 testadas por teste de raspagem. e Comparação da migração celular em células tratadas com sh-CCAT1 ou sh-SOX2. f Detecção da capacidade de invasão de células em células tratadas com sh-CCAT1 ou sh-SOX2 pelo ensaio Transwell. g Comparação da capacidade de invasão de células tratadas com sh-CCAT1 ou sh-SOX2. h O experimento de formação de esferas testou a capacidade de autorrenovação em células tratadas com sh-CCAT1 ou sh-SOX2. eu Comparação da taxa de formação de esfera em células tratadas com sh-CCAT1 ou sh-SOX2. a p <0,05 vs. o grupo sh-SOX2 NC. b p <0,05 vs. o grupo sh-CCAT1 NC. Os dados de medição foram descritos como média ± desvio padrão, e as comparações entre vários grupos foram avaliadas por ANOVA de uma via seguida pelo teste post hoc de Tukey
MiR-185-3p esgotado reverte o papel da regulação negativa de CCAT1 em CD44 + Células HeLa
Em seguida, examinamos se miR-185-3p estava envolvido no processo de regulação de CCAT1 da proliferação e auto-renovação de células-tronco CC. CD44 + HeLa foi transfectado com miR-185-3p mimetizador ou co-transfectado com sh-CCAT1 e inibidor miR-185-3p. Os resultados apresentaram que a regulação positiva de miR-185-3p reduziu notavelmente a proliferação, migração, invasão e taxa de formação de esferas enquanto aumentou a taxa de apoptose de CD44 + Células HeLa. As células tratadas com o inibidor miR-185-3p podem reverter o papel do CCAT1 regulado negativamente na proliferação, migração, invasão, apoptose e formação de esfera celular de CD44 + Células HeLa (Fig. 4a-i).
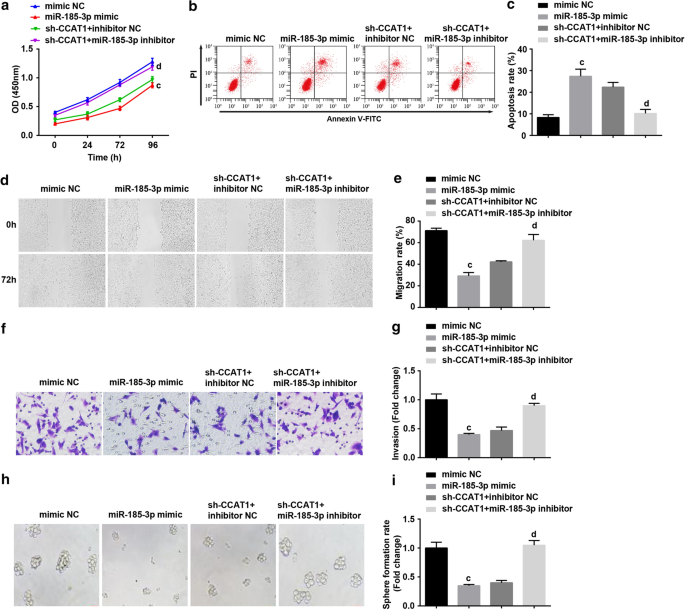
O miR-185-3p superexpresso suprime a taxa de proliferação, migração, invasão e formação de esferas e aumenta a apoptose de CD44 + Células HeLa. a O ensaio CCK-8 testou a curva de crescimento celular em células tratadas com miR-185-3p mimetizador. b A citometria de fluxo detectou apoptose celular em células tratadas com miR-185-3p mimetizador. c Comparação da taxa de apoptose celular em células tratadas com miR-185-3p mimetizador. d Migração de células em células tratadas com miR-185-3p mimetizado testado por teste de raspagem. e Comparação da migração celular em células tratadas com miR-185-3p mimetizador. f Detecção da capacidade de invasão de células tratadas com miR-185-3p mimetizador por ensaio Transwell. g Comparação da capacidade de invasão de células tratadas com miR-185-3p mimetizador. h O experimento de formação de esferas testou a capacidade de autorrenovação em células tratadas com miR-185-3p mimetizador. eu Comparação da taxa de formação de esfera em células tratadas com miR-185-3p mimetizador. a p <0,05 vs. o grupo sh-SOX2 NC. b p <0,05 vs. o grupo sh-CCAT1 NC. c p <0,05 vs. o grupo NC mímico. d p <0,05 vs. grupo NC do inibidor sh-CCAT1 +. Os dados de medição foram descritos como média ± desvio padrão, e as comparações entre vários grupos foram avaliadas por ANOVA de uma via seguida pelo teste post hoc de Tukey
Lowly Expressed SOX2 e Lowly Expressed CCAT1 diminuem a expressão FOXP3 e aumentam a expressão miR-185-3p no CD44 + Células HeLa
Posteriormente, examinamos a expressão de SOX2 / CCAT1 / miR-185-3p / FOXP3 em CD44 + As células HeLa após a transfecção de sh-SOX2, sh-CCAT1, miR-185-3p mimetizam e co-transfecção de sh-CCAT1 e inibidor de miR-185-3p. As expressões de SOX2, CCAT1 e FOXP3 foram reduzidas, enquanto a expressão de miR-185-3p foi elevada em células tratadas com sh-SOX2. As expressões de CCAT1 e FOXP3 foram reduzidas e a expressão de miR-185-3p foi aumentada em células tratadas com sh-CCAT1. A expressão de miR-185-3p foi elevada e a expressão de FOXP3 diminuiu em células introduzidas com o mimetizador de miR-185-3p. A expressão de FOXP3 foi elevada e a expressão de miR-185-3p foi reduzida em células transfectadas sucessivamente com sh-CCAT1 e inibidor de miR-185-3p (Fig. 5a-d).
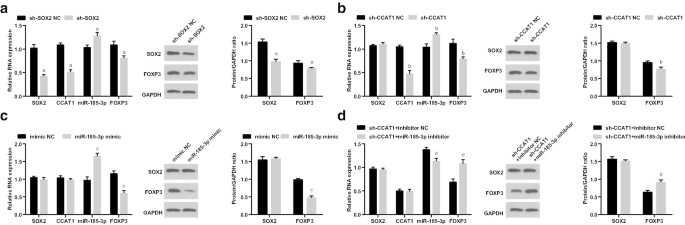
SOX2 pouco expresso e CCAT1 pouco expresso diminuem a expressão de FOXP3 e aumentam a expressão de miR-185-3p em CD44 + Células HeLa. a Expressão de SOX2 / CCAT1 / miR-185-3p / FOXP3 nos grupos sh-SOX2 NC e sh-SOX2. b Expressão de SOX2 / CCAT1 / miR-185-3p / FOXP3 nos grupos sh-CCAT1 NC e sh-CCAT1. c Expressão de SOX2 / CCAT1 / miR-185-3p / FOXP3 nos grupos mímicos NC e miR-185-3p mímico. d Expressão de SOX2 / CCAT1 / miR-185-3p / FOXP3 nos grupos de inibidor sh-CCAT1 + NC e sh-CCAT1 + miR-185-3p inibidor. a p <0,05 vs. o grupo sh-SOX2 NC. b p <0,05 vs. o grupo sh-CCAT1 NC. c p <0,05 vs. o grupo NC mímico. d p <0,05 vs. grupo NC do inibidor sh-CCAT1 +. Os dados de medição foram descritos como média ± desvio padrão, e as comparações entre vários grupos foram avaliadas por ANOVA de uma via seguida pelo teste post hoc de Tukey
SOX2 liga-se a CCAT1 que afeta a expressão de miR-185-3p e FOXP3 é um gene alvo de miR-185-3p
Os locais de ligação potencial do fator de transcrição da região do promotor CCAT1 foram predicados e analisados pelo site https://jaspar.genereg.net/, e mostrou que SOX2 e CCAT1 tinham locais de ligação potenciais na região do promotor CCAT1 (Fig. 6a). ChIP-qPCR relatou que (Fig. 6b):Em contraste com o grupo IgG, mais fragmentos do promotor CCAT1 foram enriquecidos no grupo SOX2 no local de ligação E1, o que provou que SOX2 estava ligado ao promotor CCAT1 no local E1 e SOX2 estava envolvidos na regulamentação do CCAT1. O ensaio do gene repórter de luciferase dupla exibiu que (Fig. 6c):A atividade da luciferase foi suprimida em células co-transfectadas com sh-SOX2 e E1-WT, indicando que SOX2 poderia se ligar a CCAT1.
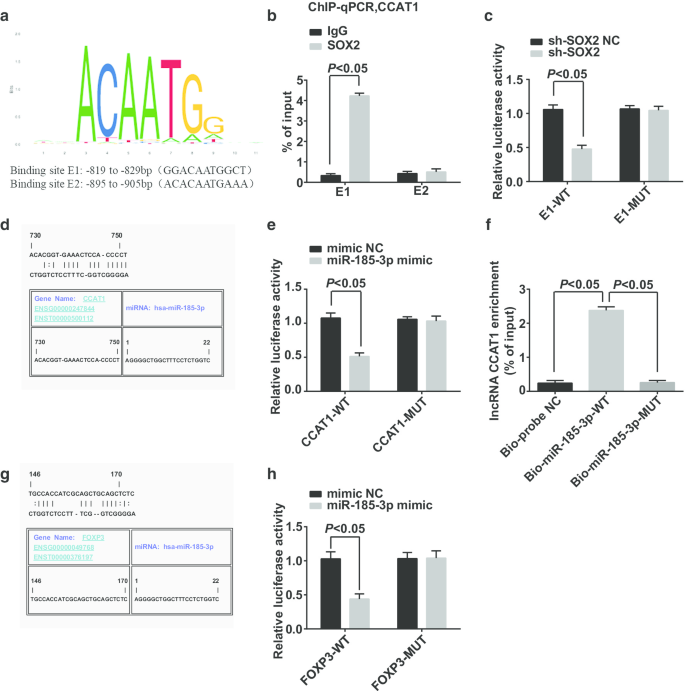
SOX2 pode se ligar a CCAT1 que afeta a expressão de miR-185-3p, e FOXP3 é um gene alvo de miR-185-3p. a Predição de sítios de ligação em regiões promotoras SOX2 e CCAT1 por sítios de bioinformática. b O experimento ChIP-qPCR verificou a relação de ligação entre SOX2 e CCAT1. c O sítio de ligação de SOX2 e CCAT1 verificado por ensaio de gene repórter dual luciferase. d Predição de sítios de ligação em CCAT1 e miR-185-3p por sítios de bioinformática. e Verificação da ligação de CCAT1 e miR-185-3p por ensaio de gene repórter de luciferase dupla. f A relação de ligação entre CCAT1 e miR-185-3p em células verificada por ensaio pull-down de RNA. g Previsão da relação de segmentação entre miR-185-3p e FOXP3 pelo site de bioinformática. h Identificação da relação de direcionamento entre miR-185-3p e FOXP3 por ensaio de gene repórter de luciferase duplo. Os dados de medição foram representados como média ± desvio padrão, e as comparações entre dois grupos foram avaliadas pelo teste t
O site de Jefferson previu que CCAT1 poderia se ligar a miR-185-3p (Fig. 6d). O ensaio do gene repórter de luciferase duplo relatou que (Fig. 6e) a atividade da luciferase foi diminuída em células introduzidas com miR-185-3p mimetizador e CCAT1-WT, sugerindo que miR-185-3p poderia se ligar a CCAT1. O ensaio pull-down de RNA foi utilizado para verificar se CCAT1 poderia se ligar ao miR-185-3p. Os resultados revelaram que (Fig. 6f) o nível de enriquecimento de CCAT1 em células tratadas com Bio-miR-185-3p-WT aumentou acentuadamente, enquanto o nível de enriquecimento de CCAT1 em células tratadas com Bio-miR-185-3p-MUT mostrou nenhuma diferença significativa. Este resultado demonstrou que CCAT1 pode adsorver miR-185-3p, afetando assim a expressão de miR-185-3p.
A relação de destino entre miR-185-3p e FOXP3 foi prevista pelo site de Jefferson (Fig. 6g). O ensaio do gene repórter de luciferase dupla verificou que (Fig. 6h) a atividade relativa da luciferase das células reduziu drasticamente após FOXP3-WT e miR-185-3p mimetizarem co-transfectados para CD44 + As células HeLa, enquanto o FOXP3-MUT co-transfectado com miR-185-3p mimetizador não afetou a atividade relativa da luciferase das células, sugerindo que o miR-185-3p tinha como alvo o FOXP3.
Discussão
O CC é a quarta neoplasia maligna mais frequente em mulheres no mundo, seguido pelos cânceres de mama, cólon e pulmão [3]. Foi relatado que as células CC com expressão positiva de SOX2 mostram as características das células-tronco cancerosas [21]. Um estudo relatou que o CCAT1 é um lncRNA oncogênico essencial relacionado ao CC e exerce um papel facilitador no crescimento e na invasão de células CC [11]. Outro estudo revela que o miR-185-3p pode prever a radiossensibilidade do carcinoma nasofaríngeo e regular o crescimento e a apoptose das células cancerosas [22]. Foi relatado que o autoanticorpo circulante para FOXP3 reflete o progresso contínuo da lesão cervical e pode ser um biomarcador potencial para o prognóstico precoce de CC [23]. O estudo atual foi projetado para explorar como o eixo SOX2 / CCAT1 / miR-185-3p / FOXP3 influenciou a proliferação e a capacidade de auto-renovação das células-tronco CC.
Com base em nossas descobertas, as expressões de SOX2 e CCAT1 foram aumentadas em tecidos e células CC que foram associados ao tamanho do tumor, LNM e FIGO avançado. Funcionalmente comprovado, a regulação negativa de SOX2 e CCAT1 diminuiu a proliferação, migração, invasão e número de células esféricas e aumentou a apoptose de células-tronco CC. Semelhante ao nosso estudo, a expressão de SOX2 tende a aumentar em CC [24, 25]. Moreover, SOX2 expression is also up-regulated in CC cells derived from cancer stem cells [26]. Overexpressed SOX2 was suggested to link with clinicopathological characteristics of patients with several types of cancer, not limited to CC. For example, it was suggested that up-regulated SOX2 shows in cervical squamous cell carcinoma patients staged in FIGO I-II [27]. Moreover, SOX2 expression is linked to LNM in oral squamous cell carcinoma [28]. When it comes to the molecular function of SOX2 for cancer progression, there is an observational work presenting that down-regulated SOX-2 suppresses cell migration and invasion of cervical squamous cell carcinoma [29]. Meanwhile, another research has offered a proof that up-regulated SOX2 enhances CC cell clonogenicity, proliferation and tumorigenicity in vitro and in vivo than control cells [30].
Concerning to the regulatory relation between SOX2 and CCAT1, an existed study has presented that silencing SOX2 markedly reduces CCAT1 mRNA level [31]. As to the role of CCAT1 in cancers, a study has showed that CCAT1 expression is markedly elevated in CC tissues versus in the adjacent normal tissues [11, 12]. Of note, CCAT1 overexpression in CC is positively related to the tumor size [12]. In terms of the role of CCAT1 in cancer cell activity, there is a research highlighting that overexpressed CCAT1 accelerates CC cell proliferation, colony formation and invasion [11]. Interestingly, a previous research has demonstrated that the cell viability, invasive and migratory abilities are declined via knocking down CCAT1 [12]. Anyway, the functional effect of SOX2 and CCAT1 in other cancers was similar to that in CC.
Afterward, our research revealed that CCAT1 could bind to miR-185-3p, the down-regulated CCAT1 in CC and overexpressing miR-185-3p suppressed the proliferation and self-renewal abilities of CC stem cells. It is reported that CCAT1 and miR-185-3p are negatively correlated [13]. Furthermore, a result reported that a reduction is seen in miR-185-3p expression in radioresistant nasopharyngeal carcinoma cases [22]. Regarding to the suppressive function of miR-185-3p in cancer cell aggressiveness, a study has revealed that up-regulation of miR-185-3p suppresses the invasive and metastatic properties of nasopharyngeal carcinoma cells [32]. Furthermore, Zou et al. have suggested that restored miR-185 represses breast cancer cell growth and invasion [33]. There is a article finding that up-regulation of miR-185 declines the proliferation, invasion and colony formation capacities of non-small cell lung cancer cells in vitro [34]. It is presented that in vitro cell proliferation, invasion and migration as well as in vivo tumor growth are suppressed via miR-185-overexpressing in non-small cell lung cancer cells [35]. From those studies, the anti-tumor role of miR-185-3p in the present study was consistent with previous researches.
To proceed, we unveiled that miR-185-3p targeted FOXP3, the overexpressed gene in CC to regulate CC stem cell activities. In fact, FOXP3, the regulator of SOX2 cancer stem-like cell marker in colon cancer [36], has been investigated in CC, showing an up-regulation in CC cells [19] [37]. It was evidenced that elevating FOXP3 promotes the formation of tumor spheres and stimulates the stemness of non-small cell lung cancer cells [38].
Conclusion
Collectively, we explored for the first time that SOX2 transcription could activate CCAT1, thereby inhibiting miR-185-3p and regulating FOXP3 to promote the proliferation and self-renewal of CC stem cells, which is a potential avenue to treat CC. Additionally, however, limitations in this present study still exist in the relatively small trial size in the designed experiment. Thus, clinical researches might be further carried out to detect the efficacy for the treatment of CC.
Disponibilidade de dados e materiais
Não aplicável.
MicroRNA-301 inibido restringe a angiogênese e o crescimento celular no carcinoma de células escamosas do esôfago por elevação do PTEN
Aplicação multifuncional de nanocompósito de óxido acoplado de Zn-Fe-Mn auxiliado por PVA
Nanomateriais
- Demonstração de um biossensor flexível à base de grafeno para detecção sensível e rápida de células cancerosas do ovário
- Nanopartículas para terapia do câncer:progresso e desafios atuais
- Administração de medicamentos baseados em células para solicitações de câncer
- Nanotécnicas Inativam células-tronco cancerosas
- Efeito da distribuição de nanopartículas de ouro em TiO2 nas características ópticas e elétricas de células solares sensibilizadas por corante
- Óxido de grafeno hibridizado nHAC / PLGA Scaffolds facilitam a proliferação de células MC3T3-E1
- Estudos de TEM e STEM nas morfologias transversais de filmes anti-reflexo de SiO2 de banda larga dupla / tripla
- Fabricação e caracterização de Nano-Clips de ZnO pelo processo mediado por poliol
- Dieletroforese de ondas moduladas por frequência de vesículas e células:inversões periódicas na frequência de cruzamento
- Fatores que afetam o sucesso e a segurança do processo de puncionamento