


Papéis de ROS e parada do ciclo celular na genotoxicidade induzida pela nanoestrutura de núcleo de ouro nanorod / casca de prata
Resumo
Para entender a genotoxicidade induzida no fígado por nanopartículas de prata (AgNPs) e íons de prata, um núcleo nanobastão de ouro / nanoestrutura de concha de prata (Au @ Ag NR) e células HepaRG de hepatócitos humanizados foram usados neste estudo. O envolvimento de estresse oxidativo e parada do ciclo celular no DNA e dano cromossômico induzido por 0,4–20 µg mL −1 Au @ Ag NR foram investigados pelo ensaio do cometa, ensaio γ-H2AX e teste do micronúcleo. Além disso, a distribuição de Au @ Ag NR foi analisada. Nossos resultados demonstraram que tanto Ag + e Au @ Ag NR levou à clivagem de DNA e danos cromossômicos (clastogenicidade) em células HepaRG e que o Au @ Ag NR retido no núcleo pode liberar Ag + , agravando os danos, que são causados principalmente pela parada do ciclo celular e formação de ROS. Os resultados revelam a correlação entre o acúmulo intracelular, Ag + liberação de íons e potencial genotoxicidade de AgNPs.
Introdução
Nanopartículas de prata (AgNPs), de 1 a 100 nm de tamanho, podem exibir um amplo espectro de propriedades antimicrobianas, penetrando patógenos e inativando o grupo sulfidrila interno de suas enzimas metabólicas [1]. Eles demonstraram bacteriostase potente e efeitos bactericidas em Escherichia Coli , Neisseria gonorrhoeae e Chlamydia trachomatis , e são amplamente utilizados como revestimentos médicos, produtos domésticos [2] e também curativos para feridas [3]. Evidências convincentes mostram que as nanopartículas são capazes de entrar no núcleo e interferir no processo de síntese e transcrição do DNA [4]. Em nosso estudo anterior, relatamos que uma única dose intravenosa de 5 mg / kg de AgNPs poderia introduzir notável quebra de cromossomos nas células da medula óssea de ratos Sprague-Dawley [5]. Uma única injeção intraperitoneal de 10 mg / kg ou mais de AgNPs induziu danos ao DNA e ao cromossomo [6]. Flower et al. [7] sugeriu que AgNPs em doses de 50 e 100 μg mL −1 pode desencadear danos ao DNA cinco minutos após a administração, destacando a genotoxicidade da prata (Ag) de liberação rápida. Considerando o risco de exposição excessiva, a investigação da NanoGenotoxicologia ou do dano ao DNA e potencial carcinogênico dos nanomateriais projetados tem recebido muita atenção [8].
Os principais mecanismos para lesões genéticas induzidas por AgNP são considerados a superprodução de espécies oxidativas reativas, inflamação e distúrbio do ciclo celular [9, 10]. Como sugerido em estudos anteriores, os AgNPs podem interagir diretamente com o DNA via dano oxidativo [11] e interferir na interfase no nível do DNA e na mitose no nível cromossômico, ou interagir com a nucleoproteína e o aparelho do fuso mitótico para perturbar os pontos de verificação do ciclo celular [ 12]. No entanto, se a genotoxicidade induzida por AgNPs é parcialmente atribuível às nanopartículas [13, 14] ou completamente ao Ag + liberado íons ainda não está claro [15, 16].
Investigar a genotoxicidade dos AgNPs é difícil devido à liberação instável e ininterrupta da prata nos tecidos, dificultando a localização dos AgNPs e a diferenciação do nanocore do Ag. Nosso grupo desenvolveu recentemente uma nanoestrutura de núcleo de nanobastão de ouro / casca de prata (Au @ Ag NR) para estudar a toxicidade induzida por nanopartículas [17]. O núcleo de ouro do Au @ Ag NR é fisiologicamente inato no tecido e pode ser usado como um padrão interno para monitorar a liberação de Ag + íons da haste monitorando a mudança na razão Ag / Au, medida usando espectrometria de massa com plasma acoplado indutivamente (ICP-MS) [18]. Por este método, as diferentes origens das toxicidades podem ser identificadas. Estudos anteriores mostraram que o Ag + lançado Os íons da casca do Au @ Ag NR resultaram em dano oxidativo aos rins e, eventualmente, levaram a mudanças morfológicas e comprometimento da função de filtração do glomérulo [19]. Jiang et al. [20] sugeriram que tanto a atividade específica da partícula quanto a liberação intracelular do íon prata pelo Au @ Ag NR contribuem para a resposta tóxica das células da granulosa. Também adotamos Au @ Ag NR como modelo para estudar o potencial de genotoxicidade in vivo de AgNPs e demonstramos que a clastogenicidade, e não a mutagenicidade, é a forma primária de genotoxicidade induzida tanto pela casca de Ag quanto pelo Ag + íons, enquanto não houve diferença em seus padrões de toxicidade [21].
O fígado é um dos principais órgãos com tendência ao acúmulo de AgNPs e é reconhecido como órgão / tecido alvo para a genotoxicidade induzida por AgNPs. Nosso estudo anterior mostrou que alguma quantidade de prata (8,26 ± 3,90 μg / g) e ouro (80,07 ± 64,72 μg / g) permaneceu no fígado de ratos SD oito semanas após a administração intravenosa de uma dose de Au @ Ag NR [21 ] Neste estudo, tentamos identificar os papéis da parada do ciclo celular e do estresse oxidativo reativo no cromossomo induzido por AgNP e danos no DNA usando Au @ Ag NR em células HepaRG derivadas de hepatoma humano. Ensaios de genotoxicidade, incluindo ensaio cometa, ensaio γ-H2AX e teste de micronúcleo, foram realizados em paralelo com o sequestrador de radicais oxidativos para investigar a contribuição de espécies reativas de oxigênio (ROS) em danos ao DNA / cromossomo, enquanto a apoptose celular, ciclo celular e proteínas relacionadas foram determinados para explorar os mecanismos pelos quais AgNPs interrompem a síntese e replicação de DNA. Além disso, a acumulação e distribuição intracelular de Au @ Ag NR foi investigada combinando espectrometria de massa de plasma acoplado indutivamente (ICP-MS) e microscopia eletrônica de transmissão (TEM) para diferenciar o papel das nanopartículas e íons de Ag liberados.
Materiais e métodos
Cultura de células e tratamento
A linha celular de hepatoma humano HepaRG (Thermo Fisher Scientific) foi usada neste estudo. As células foram cultivadas em RPMI 1640 contendo 10% de soro fetal bovino (FBS, Australia Origin, Gibco) e 1% de solução de penicilina-estreptomicina-glutamina (Gibco) em uma atmosfera umidificada de 5% de CO 2 a 37 ° C. As células foram tratadas com concentrações crescentes de Au @ Ag NR por 24 h ou 72 h, respectivamente, e as concentrações foram determinadas de acordo com IC 50 estimado por ensaio de viabilidade celular. Para investigar o papel de ROS na genotoxicidade, 1 mM N -Acetil-l-cisteína (NAC, Sigma-Aldrich) foi aplicada durante 1 h antes do tratamento com Au @ Ag NR.
Ensaio de crescimento / viabilidade celular ATP
As células foram semeadas em uma placa de 96 poços a uma densidade de 5 × 10 3 /Nós vamos. Após 24 h de incubação, o meio foi aspirado e as células foram expostas a diferentes concentrações de Au @ Ag NR por 24 h ou 48 h, respectivamente. Um amplo espectro de concentrações foi preparado e quatro poços por tratamento foram realizados em um período de tratamento. A citotoxicidade do Au @ Ag NR foi examinada pelo ensaio de trifosfato de adenosina (ATP) (CellTiter-Glo® 2.0 Assay, Promega), que mede a atividade metabólica celular quantificando a quantidade de ATP, um importante parâmetro do metabolismo em células viáveis. Os sinais luminescentes, que refletem a quantidade de células viáveis, foram detectados usando VICTOR Multilabel Plate Reader (2030-0050, PerkinElmer) e IC 50 os valores foram estimados como a concentração de Au @ Ag NR para metade da viabilidade máxima por Prism 7 (GraphPad Prism 7, CA, EUA). A razão de viabilidade é calculada usando a seguinte equação:
$$ {\ text {Viabilidade}} \, {\ text {Ratio}} \, \ left (\% \ right) ={\ text {RLU}} _ {{{\ text {sample}}}} / { \ text {RLU}} _ {{{\ text {veículo}}}} \ vezes {1} 00 \% $$
onde RLU é a unidade de luz relativa representada como o valor médio de quatro poços, RLU veículo células representadas não tratadas com nanobastões e RLU amostra células representadas que foram tratadas com diferentes concentrações de Au @ Ag NR.
Determinação da concentração de prata e ouro nas células
As amostras de células foram digeridas em ácido nítrico usando o sistema de digestão por micro-ondas. Após a digestão, as amostras foram preparadas com uma mistura contendo ácido nítrico 1% e ácido clorídrico. As quantidades de Ag e Au nas soluções foram determinadas por ICP-MS (NexION300X, PerkinElmer). A análise de TEM foi usada para determinar a presença de Au NR e Au @ Ag NR na célula. As amostras de células foram fixadas em uma mistura de glutaraldeído a 2,5% e paraformaldeído a 2% por 2h a 4 ° C. Os sedimentos celulares foram fixados e lavados três vezes em tampão fosfato (pH 7,4) e pós-fixados em tetróxido de ósmio a 1% por 2 h a 4 ° C. As amostras foram posteriormente enxaguadas em água destilada três vezes e desidratadas por 15 min em diferentes concentrações de etanol (50%, 70%, 90% e 100% de etanol, respectivamente) uma após a outra. Posteriormente, o óxido de propileno em diluições de 1:1 e 1:3 foi aplicado à resina a 20–26 ° C por 2 h. A polimerização foi realizada por aquecimento gradual a 35 ° C durante 16 h, 45 ° C durante 8 h, 55 ° C durante 14 h e 65 ° C durante 48 h. Cortes ultrafinos foram corados por 25 min com acetato de uranila e citrato de chumbo e analisados por um microscópio eletrônico de transmissão (H-7650, HITACHI, Japão).
Ensaio cometa convencional e modificado
As células foram semeadas em placas de 12 poços em densidades de 2 × 10 5 / well ou 3 × 10 5 / poço para um tratamento de 24 ou 72 h, respectivamente. Peróxido de hidrogênio (H 2 O 2 ) a uma concentração de 200 μmol foi exposto às células como controle positivo por uma hora. Para cada amostra, dois poços foram preparados tanto para o tratamento convencional quanto para o tratamento com formamidopirimidina glicosilase (Fpg). O ensaio convencional do cometa foi realizado em condições alcalinas (pH> 13), conforme descrito anteriormente [21]. Para os poços tratados com Fpg, um tratamento Fpg adicional foi aplicado antes do procedimento de desenrolamento do DNA, e as lâminas foram imersas em um tampão de enzima (0,1 M KCl, 0,5 mM EDTA, 40 mM HEPES, 0,2 mg.mL −1 BSA) três vezes por 5 min cada. O Fpg (New England Biolabs, Inc., UK) foi diluído a 1:50.000 com tampão de enzima. Alíquotas de cem mililitros da enzima diluída foram adicionadas a cada gel nas lâminas de microscópio e incubadas em uma câmara de umidade a 37 ° C por 30 min. As demais etapas foram as mesmas do tratamento convencional. Os ensaios do cometa foram realizados em triplicado. Pelo menos 50 células por amostra foram avaliadas de forma independente usando o microscópio fluorescente Nikon Eclipse 80i (Nikon, Tóquio, Japão), enquanto Komet 6.0 (Andor Technology, Belfast, Reino Unido) foi usado para analisar o valor médio da porcentagem de DNA na cauda e cauda verde-oliva momento (OTM) de cada amostra.
Qualificação de focos γ-H2AX por citometria de fluxo e triagem de alto conteúdo
Para a quantificação por citometria de fluxo, as células foram semeadas em placas de 12 poços em densidades de 2 × 10 5 / well ou 3 × 10 5 / poço para um tratamento de 24 ou 72 h, respectivamente, enquanto para o ensaio de triagem de alto conteúdo, as células foram semeadas em placas de 96 poços em densidades de 6 × 10 3 / well ou 1 × 10 4 / poço para um tratamento de 24 ou 72 h, respectivamente. Como controle positivo, 2 μM de metanossulfonato de metila (MMS, Sigma-Aldrich) foi aplicado em paralelo com as células por uma hora. As células foram lavadas em solução salina tamponada com tris (TBS) e fixadas com paraformaldeído a 4% por 15 min em temperatura ambiente. Após a lavagem com TBS, as células foram incubadas com 50 μL de metanol gelado por 30 min a -20 ° C. As células foram posteriormente enxaguadas em TBS três vezes e o reagente de bloqueio (TBS contendo 0,3% de Triton X-100 e 10% de soro de cabra) foi aplicado durante 1 h. O anticorpo primário (anti-fosfo-H2AX Ser139 de camundongo, Millipore) foi diluído a 1:200 com reagente de bloqueio e incubado com as células durante a noite a 4 ° C. A placa foi então novamente enxaguada com TBS por três vezes, e o anticorpo secundário (Alexa Fluor 488 cabra anti-camundongo, Life Technologies), diluído com o reagente de bloqueio na proporção de 1:20, foi adicionado subsequentemente. As amostras foram mantidas no escuro em temperatura ambiente por 1 h e 2 μg mL −1 (20 μL / poço) DAPI (Invitrogen) foi adicionado a cada poço. A fluorescência foi medida usando citometria de fluxo (FACSCalibur, BD Bioscience, NJ, EUA) ou High Content Analysis System (Operetta CLS, PerkinElmer). Para o ensaio de citometria de fluxo, foram analisados dados de pelo menos 10.000 células por grupo e os experimentos foram realizados em triplicata; para análise de alto conteúdo, 20 campos visuais em cada poço e pelo menos cinco poços em cada grupo foram analisados.
Ensaio de citocitoma de micronúcleo de bloqueio de citocinese (CBMN-cyt)
CBMN-cyt foi realizado de acordo com o procedimento descrito por Fenech et al. [22]. As células foram semeadas em placas de 12 poços em densidades de 2 × 10 5 / well ou 3 × 10 5 / poço para um tratamento de 24 ou 72 h, respectivamente. 0,2 μg mL −1 Mitomicina C (MMC, Tokyo Chemical Industry Co., Ltd. Japão) foi exposta às células como controle positivo por 24 h. 3 μg mL −1 a citocalasina B foi aplicada após um tratamento de 24 ou 72 horas para bloquear o processo de citocinese, e as células foram colhidas após 40 horas. As amostras foram coradas com 5% de Giemsa após hipotonicidade com 0,075 mol L −1 pré-aquecido KCl e fixação com mistura 3:1 de metanol e ácido acético. Foram preparados poços em triplicado por grupo e examinadas pelo menos 1000 células binucleadas por poço.
Medição de MDA, GSH total e conteúdo de SOD
As células foram cultivadas em placas de 12 poços em densidades de 5 × 10 5 / well ou 3 × 10 5 / poço para um tratamento de 24 ou 72 h, respectivamente. Posteriormente, as células foram colhidas e lavadas três vezes com solução salina tampão de fosfato (PBS). As quantidades de malondialdeído (MDA) nos homogenatos celulares foram determinadas usando um método à base de ácido tiobarbitúrico (Nanjing Jiancheng Bio-engineering Institute, Nanjing, China). As quantidades de glutationa total (GSH) e superóxido dismutase (SOD) foram determinadas usando a quantificação de glutationa total e kits de ensaio SOD (Dojindo Molecular Technologies, Inc. Kumamoto, Japão), respectivamente. As densidades ópticas (O.D) de cada poço foram medidas usando VICTOR Multilabel Plate Reader (2030-0050, PerkinElmer).
Análise de citometria de fluxo para o ciclo celular
As células foram cultivadas em placas de 6 poços em densidades de 1 × 10 6 / well ou 5 × 10 5 / poço para um tratamento de 24 ou 72 h, respectivamente, e foram posteriormente fixados com etanol 70% a 4 ° C durante a noite. As amostras foram enxaguadas três vezes com PBS e coradas com tampão de coloração PI / Rnase (BD Biosciences) durante 15 min à temperatura ambiente. Populações de células sob as fases G0 / G1, S e G2 / M entre 20.000 células foram determinadas empregando regiões com área FL2 versus largura FL2. A análise foi feita por citometria de fluxo (FACSCalibur, BD Bioscience, NJ, EUA) e FlowJo (BD Bioscience), e os experimentos foram realizados em triplicata.
Análise de citometria de fluxo de apoptose celular
As células foram cultivadas em placas de 6 poços em densidades de 1 × 10 6 / well ou 5 × 10 5 / poço para um tratamento de 24 ou 72 h, respectivamente. Posteriormente, foram lavados duas vezes com PBS e diluídos com 500 μL de tampão de ligação 1 × (FITC Annexin V Apoptosis Detection Kit I, BD Bioscience) para ajustar a suspensão para cerca de 1 × 10 6 células / mL e, subsequentemente, 100 μL de diluição foram misturados com 5 μL de FITC Anexina V e 5 μL de PI. As amostras foram coradas em temperatura ambiente por 15 min, e pelo menos 10.000 células foram analisadas para determinar a população de células em apoptose precoce e tardia, empregando regiões com FL1H versus FL2H usando citometria de fluxo (FACSCalibur, BD Bioscience, NJ, EUA) e FlowJo (BD Bioscience). Os experimentos foram realizados em triplicata.
Análise de Western blot
As células foram cultivadas em um 2 de 75 cm frasco em densidades de 1 × 10 7 / poço e 6 × 10 6 / poço para um tratamento de 24 e 72 h, respectivamente. As células foram lisadas com tampão de lise RIPA contendo inibidor de protease (PMSF), e a concentração de proteínas foi determinada usando um kit de quantificação de proteínas BCA (Beyotime Biotechnology, China). As concentrações das amostras foram ajustadas usando tampão de lise RIPA antes da desnaturação por aquecimento a 95 ° C durante 3 min. As amostras de proteínas foram separadas por eletroforese em géis de poliacrilamida SDS 12% e transferidas para membranas de nitrocelulose (Millipore). As membranas foram bloqueadas com leite desnatado a 5% por 30 min e incubadas com p53 primário (SC-137174, Santa Cruz), p21 (SC-6246, Santa Cruz) e β-actina (sc-47778, Santa Cruz) e anticorpos secundários IgG (H + L) -HRP de cabra anti-camundongo (SE131, solabio), respectivamente. Os níveis de expressão das proteínas-alvo nas amostras foram visualizados usando um método de quimioluminescência aprimorada (ECL) e analisados pelo sistema ImageJ (National Institutes of Health).
Análises estatísticas
Os dados foram apresentados como a média ± SEM. A análise de variância unilateral (ANOVA) foi usada para testar a significância estatística das diferenças entre os grupos de controle negativo e tratados, seguida pelo teste de comparação múltipla de Dunnett usando SPSS (versão 22, IBM, Armonk, NY, EUA), e os dados foram considerados estatisticamente significativo em P <0,05. As figuras foram preparadas no GraphPad Prism 7 for Windows (GraphPad Software, La Jolla, CA, EUA).
Resultados
Caracterização de Au NR e Au @ Ag NR
Nanobastões de ouro (Au NRs), núcleos de nanobastões de ouro e nanoestruturas de casca de prata (Au @ Ag NR) foram projetados, preparados e caracterizados conforme descrito anteriormente [21]. Resumidamente, os diâmetros e comprimentos médios são 15,0 ± 2,5 nm, 66,7 ± 2,5 nm para Au NRs e 26,2 ± 3,0 nm, 72,7 ± 8,9 nm para Au @ Ag NRs. A espessura da casca Ag é de cerca de 5 nm. Os potenciais zeta de Au NRs e Au @ Ag NRs dispersos em água revestidos com PDDAC foram 37,7 ± 1,6 mV e 52,5 ± 1,4 mV, respectivamente. A razão em peso Ag / Au do Au @ Ag NR preparado foi estimada em 2,3. Os resultados da caracterização são mostrados na Fig. 1.
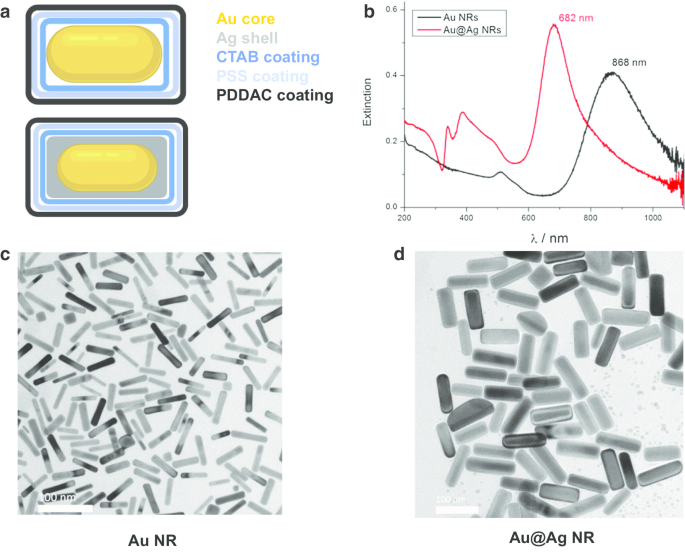
Caracterização de Au NR e Au @ Ag NR. a Diagrama estrutural de Au NR e Au @ Ag NR; b Espectros de extinção UV-Vis-NIR de Au NR e Au @ Ag NR dispersos em água; c imagens representativas de TEM de Au NR; d imagens TEM representativas de Au @Ag NR
Viabilidade celular
A citotoxicidade de Au @ Ag NR em relação às células HepaRG foi investigada pelo ensaio de viabilidade de ATP (Tabela 1), e as células foram expostas a Au @ Ag NR por 24 ou 48 h em concentrações que variam de 0,125 a 160 μg mL −1 . Au @ Ag NR induziu efeitos citotóxicos significativos de maneiras dependentes do tempo e da dose após exposição de 24 e 48 h, com% de viabilidade IC 50 a 20 µg mL −1 e 6 µg mL −1 , montado pelo software GraphPad Prism 7.0, respectivamente. Considerando a citotoxicidade geral, os períodos de tratamento foram ajustados para 24 he 72 h, enquanto as concentrações aplicadas foram determinadas em 0,8 µg mL −1 , 4 µg mL −1 e 20 µg mL −1 . Além disso, Au NR foi incluído como um controle inerte, e o conteúdo de Au no grupo AuNR foi igual a 20 µg mL −1 Au @ Ag NR, que é 16 µg mL −1 . Em contraste, o pré-tratamento de 1 mM NAC foi adotado no grupo Au @ Ag NR + NAC como um controle para a resposta ao estresse oxidativo (a concentração de Au @ Ag NR é 20 µg mL −1 )
Distribuição de células de Au NR e Au @ Ag NR
A distribuição do conteúdo de Au e Ag nas células HepaRG foi analisada por ICP-MS. Conforme mostrado nas Tabelas 2 e 3, o conteúdo de Ag aumentou de uma maneira dependente da dose. No entanto, o antioxidante N -Acetil-L-cisteína (NAC) como eliminador de radicais livres pode restringir a absorção celular de nanopartículas, visto que menor teor de Ag foi observado, embora a mesma concentração de Au @ Ag NR (20 µg mL −1 ) foi aplicado neste grupo. O declínio na razão Ag / Au de 24 para 72 h indicou uma liberação contínua de Ag + da casca de Au @ Ag NR. Além disso, a absorção celular de Ag é muito maior do que Au (Tabela 4). Além disso, os dados de TEM mostraram que a maior parte do Au NR e Au @ Ag NR foram retidos nas células como aglomerados. As estruturas dos nanobastões foram claramente vistas dentro das células sujeitas à exposição de Au NR ou Au @ Ag NR sem entrar no núcleo (Fig. 2).
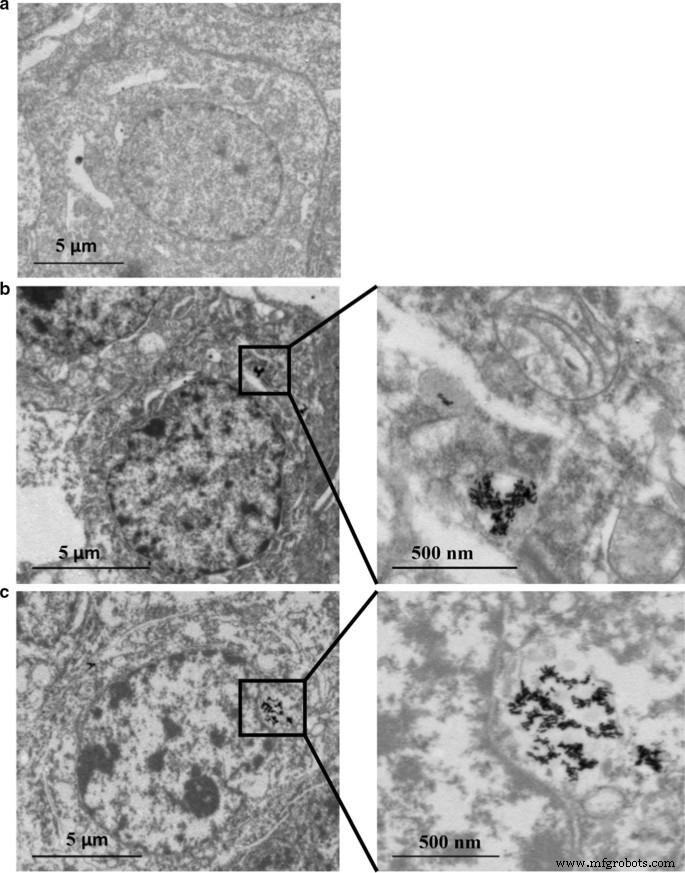
Au NR e Au @ Ag NR internalização:HepaRG por TEM a 80 kV após 24 h de exposição a 16 μg mL −1 Au NR e 20 μg mL −1 Au @ Ag NR. a Controle de veículos; b Au NR; c Au @ Ag NR
Danos no DNA
O dano ao DNA desencadeado pelo Au @ Ag NR foi avaliado tanto pelo ensaio do cometa quanto pelo ensaio γH2AX (Fig. 3). Foi observado a partir do ensaio do cometa que 0,8 a 20 µg mL −1 Au @ Ag NR pode causar danos significativos ao DNA. Depois de uma exposição de 24 ou 72 horas ao Au @ Ag NR, tanto a% de DNA da cauda quanto o OTM das células aumentaram de maneira dependente do tempo e da concentração. Além disso, danos ao DNA associados à indução de estresse oxidativo foram observados nas células tratadas com 20 µg mL −1 Au @ Ag NR pelo ensaio do cometa modificado por enzima Fgp (Fig. 3a, b). Para avaliar a extensão da quebra da fita dupla que representa uma correlação mais alta com a gênese do câncer, tanto as células γ-H2AX-positivas quanto as intensidades médias de fluorescência em células γ-H2AX-positivas foram analisadas. Após uma exposição de 24 horas ao Au @ Ag NR, nenhuma diferença foi encontrada entre os grupos em células γ-H2AX-positivas. No entanto, 4 μg mL −1 O grupo Au @ Ag NR causou um aumento significativo após um tratamento de 72 horas. Aumentos significativos nas intensidades de fluorescência foram observados em todos os grupos Au @ Ag NR após 72 h em comparação com o controle do veículo (Fig. 3c-e, P <0,05).
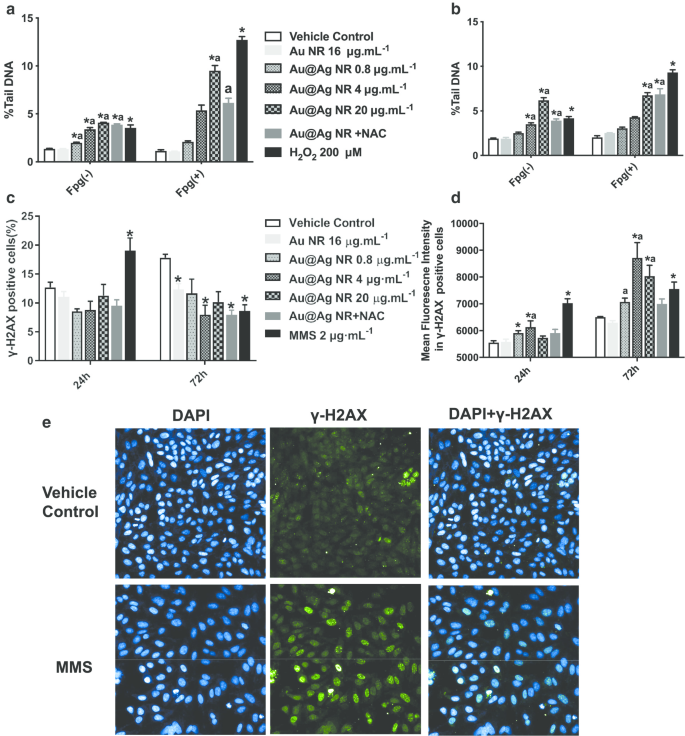
Danos no DNA induzidos por Au @ Ag NR. As células HepaGR foram expostas a Au @ Ag NR em diferentes concentrações (0,8 a 20 μg mL −1 ) por 24 he 72 h, respectivamente. a % Média de DNA de cauda após exposição a Au @ Ag NR por 24 h; b % média de DNA de cauda após exposição a Au @ Ag NR por 72 h; c porcentagem de células positivas com focos γ-H2AX estimados por citometria de fluxo; d intensidades médias de fluorescência em células com focos γ-H2AX estimados usando coloração imunofluorescente. * P <0,05 versus controle do veículo; a P <0,05 versus Au NR. 2 μM mL −1 MMS foi empregado como um controle positivo
Danos cromossômicos
A formação de micronúcleos é um biomarcador significativo para a identificação de dano cromossômico, que é um dano mais crítico ao material genético do que a quebra do DNA. A proporção de células binucleadas contendo micronúcleos foi classificada como mostrado na Fig. 4c. Au @ Ag NR aumentou a formação de micronúcleos em um padrão dependente da concentração. Após uma exposição de 24 horas, as proporções de micronúcleos observadas em células tratadas com 4 μg mL −1 Au @ Ag NR e 20 μg mL −1 Au @ Ag NR foram 1,133 ± 0,145% e 1,567 ± 0,318%, respectivamente, ambos significativamente maiores do que aqueles no grupo de controle de veículo. Após uma exposição de 72 h, a proporção de micronúcleos em células tratadas com 4 μg mL −1 Au @ Ag NR foi 1,767 ± 0,233%, o que foi significativamente maior do que o grupo de controle de veículo; a proporção de micronúcleos em células tratadas com 20 μg mL −1 Au @ Ag NR foi 2,167 ± 0,252%, o que foi significativamente maior do que aqueles observados em ambos os grupos de controle de veículo e 16 μg mL −1 Grupo Au NR (0,700 ± 0,153%). Em contraste, nenhuma diferença foi encontrada entre as células tratadas com 20 μg mL −1 Au @ Ag NR + NAC e controle de veículo, sugerindo a participação de ROS na quebra cromossômica induzida por Au @ Ag NR.
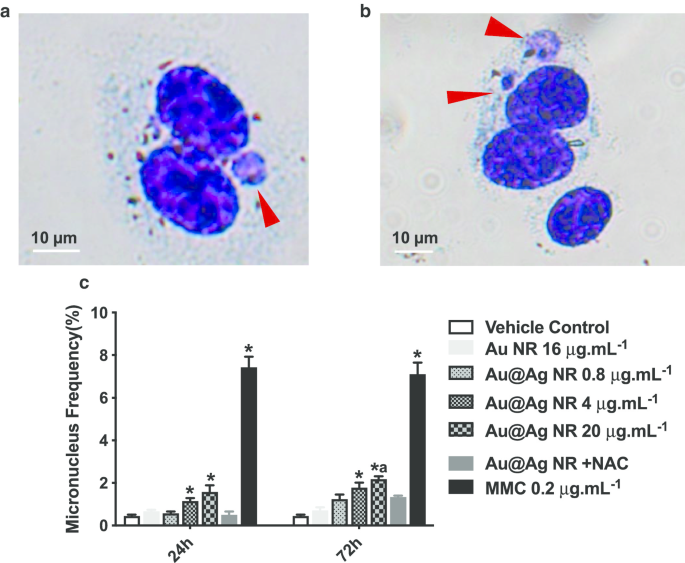
Danos cromossômicos induzidos por Au @ Ag NR. As células HepaGR foram expostas a Au @ Ag NR em diferentes concentrações de 0,8 μg mL −1 a 20 μg mL −1 por 24 he 72 h. a , b Imagens representativas do micronúcleo (seta vermelha); c frequência do micronúcleo (%). * P <0,05 versus controle do veículo; a P <0,05 versus Au NR. 0,2 μg mL −1 mitomicina C foi empregada como controle positivo
Efeitos de Au @ Ag NR na formação de ROS
Para explorar ainda mais o papel da formação de ROS no DNA induzido por Au @ Ag NR e danos cromossômicos, os níveis de MDA, GSH e SOD foram estimados. Um aumento significativo na formação de MDA ( P <0,05) foi observada após a exposição a 20 μg mL −1 Au @ Ag NR por 24 e 72 h (Fig. 5a). Além disso, os níveis de GSH e SOD em células expostas a Au @ Ag NR mostraram redução significativa ( P <0,05) de maneira dependente do tempo e da concentração. Esses resultados sugerem um desequilíbrio entre a oxidação e a antioxidação, gerado pela exposição de Au @ Ag NR (Fig. 5b, c).
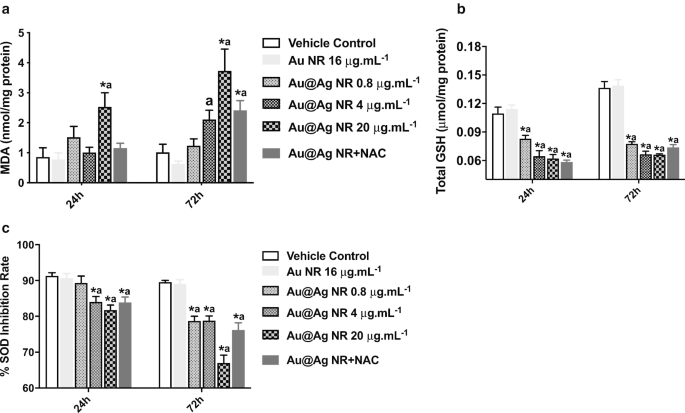
Efeitos do Au @ Ag NR na formação de ROS. As células HepaGR foram expostas a Au @ Ag NR em diferentes concentrações de 0,8 μg mL −1 a 20 μg mL −1 por 24 he 72 h. a Nível MDA; b Nível de GSH; c Nível SOD. * P <0,05 versus controle do veículo; a P <0,05 versus Au NR
Efeitos de Au @ Ag NR no ciclo celular e apoptose
Após 72 h de exposição ao Au @ Ag NR, o aumento no número de células na fase G2 / M foi observado em 4 μg mL −1 Au @ Ag NR, 20 μg mL −1 Grupo Au @ Ag NR e Au @ Ag NR + NAC, com proporções de 32,63% ± 1,77%, 32,267% ± 2,17% e 32,967% ± 4,25%, respectivamente (Fig. 6a, b), que foram significativamente maiores do que aquelas em o grupo de controle do veículo (22,37% ± 0,92%). Nesse ínterim, a apoptose celular induzida por Au @ Ag NR pode ser observada após uma exposição de 72 horas, e a taxa de apoptose tardia de células tratadas com 20 μg mL −1 Au @ Ag NR e 20 μg mL −1 Au @ Ag NR + NAC foi 78,90 ± 1,19% e 70,20 ± 4,50%, respectivamente (Fig. 6c, d). Au @ Ag NR induziu mais apoptose tardia do que apoptose precoce, e o tratamento de NAC poderia aliviar a taxa celular de apoptose tardia desencadeada por Au @ Ag NR.
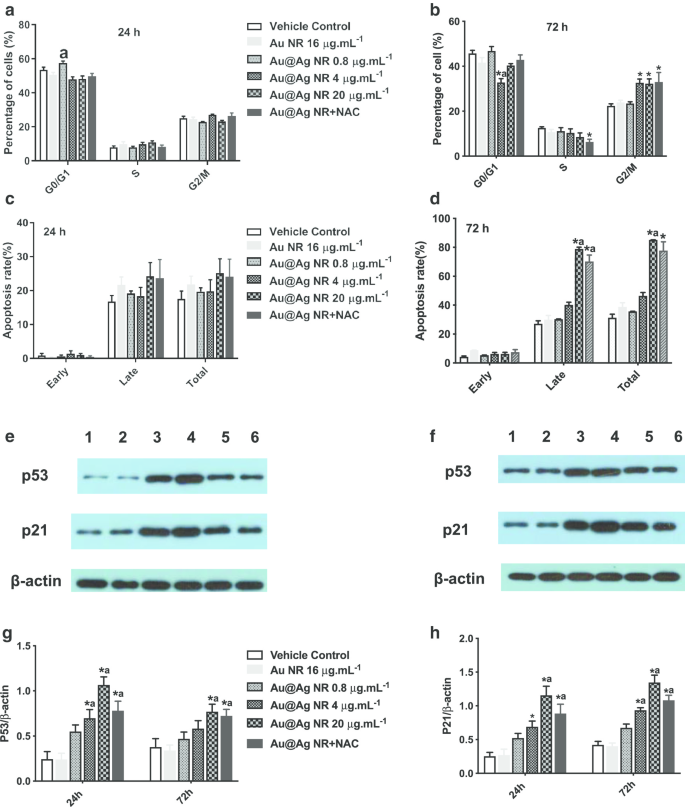
Efeitos do Au @ Ag NR no ciclo celular e na apoptose. Efeitos de Au @ Ag NR no ciclo celular ( a , b ) e apoptose ( c , d ) após exposto por 24 he 72 h, respectivamente; os dados representativos dos níveis de expressão de p53 e p21 em células HepaRG de diferentes grupos ( e , f Pista 1:controle do veículo; Pista 2:Au NR; Pista 3:Au @ Ag NR + NAC; Pista 4:Au @ Ag NR 20 μg mL −1 ; Pista 5:Au @ Ag NR 4 μg mL −1 ; Pista 6:Au @ Ag NR 0,8 μg mL −1 ); o nível de expressão relativo médio de p53 e p21 para β-actina em grupos diferentes foi resumido em ( g , f ). * P <0,05 versus controle do veículo; a P <0,05 versus Au NR
Os níveis de expressão de p21 e p53 foram detectados por Western blots e um padrão semelhante foi observado. Os níveis de expressão de p53 e p21 em células tratadas com 4 μg mL −1 e 20 μg mL −1 Au @ Ag NR aumentou significativamente ( P <0,05) e diminuíram significativamente nas células tratadas com 20 μg mL −1 Au @ Ag NR e NAC ( P <0,05, em comparação com 20 μg mL −1 Grupo Au @ Ag NR, Fig. 6e-h). Sabe-se que a proteína p53 é uma molécula central que medeia a ativação do checkpoint G2 / M em resposta a danos no DNA, e a p21 é reconhecida como um inibidor do ciclo celular dependente de p53. Assim, o Au @ Ag NR poderia interferir na replicação do DNA e dificultar o reparo do DNA pela parada do ciclo celular.
Discussão
No momento, as funções do Ag + lançado e AgNPs na geração de genotoxicidade estão longe de ser claros. Estudos anteriores de nosso grupo [21] e outros [13] demonstraram que enquanto Ag + é a principal fonte de introdução de toxicidades, as nanopartículas também podem ser altamente tóxicas. Por exemplo, AgNPs podem contribuir para a genotoxicidade induzindo a formação de radicais hidroxila [13]. Além disso, danos cromossômicos mais graves, estresse oxidativo e apoptose foram introduzidos pelo AgNP em comparação com o Ag + sozinho [23], sugerindo que diferentes vias podem estar envolvidas. Empregamos Au @ Ag NR como um material modelo para entender as formas e distribuições de AgNPs nas células, e as quantidades de Ag e Au intracelulares foram determinadas por ICP-MS. A razão em peso Ag / Au do Au @ Ag NR preparado foi estimada em 2,3. However, after a 24-h exposure, it sharply increased to 16.5 in the cells treated with Au@Ag NR, suggesting that large amount of Ag was released from the shell of Au@Ag NR within that period. When the exposure period of Au@Ag NR was extended to 72 h, the Au/Ag weight ratio was decreased to 1.7, indicating that the Ag + was released from the cell and the nanorod was the major form of Au@Ag NR in the cell at that stage. Therefore, it could be deduced that once the Au@Ag NR entered the cell, Ag + rapidly dissolved from its shell within 24 h and gradually released to the extracellular environment, while the Au@Ag NR itself retained in the cell for a longer period.
Oxidative stress is deemed as one of the most important toxicological mechanisms of nanoparticles [24]. N-acetylcysteine (NAC) is a thiol, a mucolytic agent and precursor of l-cysteine which reduced glutathione. NAC is also a source of sulfhydryl groups in cells and exerts the ROS scavenger activity by interacting with OH · e H 2 O 2 [25]. In this study, the GSH and SOD levels were significantly decreased after exposure to Au@Ag NR, while the MDA level increased in a concentration- and time-dependent manner, indicating that the Au@Ag NR introduced the oxidative stress in the cells.
The potentials of Ag and Au@Ag NR in interfering with the genetic materials were further investigated by a series of genotoxicity assays. It is noteworthy that co-culturing the NAC with Au@Ag NR could ameliorate the ROS formation, which in turn supports the participation of oxidative stress in the genotoxicity triggered by Au@Ag NR. In this study, comet and γ-H2AX assays were performed to confirm that Au@Ag NR could interact with DNA and induce certain DNA damage, and the repair endonuclease Fpg was included in the comet assay to identify the oxidative DNA damage [26]. The Fgp could recognize oxidized pyrimidines and remove oxidized purines, e.g., 8-hydroguanine, so as to create apurinic or apyrimidinic sites that could introduce gaps in the DNA strands. The oxidative stress-induced DNA breakage could be determined subsequently by another comet assay [27]. The further DNA breakage detected by the additional Fgp in the comet assay suggested that the Au@Ag NR could cause DNA damage. Mei et al. [28] observed that 5-nm-sized AgNPs induced oxidative lesion-specific DNA damage by employing the hOGG1, EndoIII and Fpg endonucleases in the comet assay. Li et al. [29] also suggested that both PVP- and silica-coated AgNPs (15–100 nm and 10–80 nm, respectively) could lead to a significant increase in DNA breakage in mice hepatocytes in the presence of hOGG1and EndoIII. The formation of γ-H2AX foci, which represents an early cellular response to genotoxic stress, is the most sensitive and specific biomarker for detecting DSBs [30]. As demonstrated in this study, γ-H2AX foci in cells exposed to Au@Ag NR were markedly increased after 24 h, and a further increase could be observed after 72 h. The reduction in the 20 µg mL −1 group might be due to the cytotoxicity to the HepaRG cells at higher concentration. Similar results were observed for AgNPs with different coatings [31, 32]. Further, our results suggest that Au@Ag NR could induce chromosome damage in HepaRG cells, as the micronucleus rates were significantly increased. This is consistent with previous studies, where AgNPs-induced increased micronucleus rate was reported in HaCaT and TK6 cells [33]. In contrast, the addition of oxidative radical scavenger NAC could inhibit the formation of micronucleus induced by Au@Ag NR. Taken together, these data suggest the participation of oxidative stress in AgNP-introduced clastogenicity risk in vitro.
Previous studies have investigated the cell cycle arrest and cytotoxicity induced by AgNPs [33,34,35]. With prolonging the exposure time, the impact of AgNPs on cell cycle and apoptosis might be enhanced and in turn aggravate the cytotoxicity and genotoxicity. Usually, the cell cycle checkpoints (e.g., G2/M) were initiated by cells when experiencing DNA damage, and this mechanism serves to prevent the cell from entering mitosis (M phase). The G2/M cell cycle arrest indicates that an increasing percentage of cells is hindered in G2 phase for DNA repairing. Cells experiencing successful DNA repairing would further proceed to mitosis; however, for those with fatal damages, irreversible G2/M cell cycle arrest and cells apoptosis would take place [36]. We observed that Au@Ag NR could arrest the majority of HepaRG cells in G2/M phase, induce late cell apoptosis and increase the expression levels of p53 and p21, which are important proteins associated with the regulation of cell cycles [37]. As p53 could also induce apoptosis, when the DNA cannot be repaired properly [38], the p21 might indirectly participate in cell apoptosis by cell cycle arrest in a p53-dependent pathway via down-regulating the nuclear protein ICBP90 for DNA replication and cell cycle regulation [39]. Furthermore, apoptosis and a G2/M arrest induced by activation of the p53/p21 system have been reported in HepG2 cells following the administration of garlic extracts [40]. Thus, it could be inferred that the oxidative stress-triggered DNA/chromosome damages might facilitate the expression of p53 and p21, which subsequently induces cell cycle arrest. Extending the exposure period of Au@Ag NRs to the DNA/chromosome during replication may further aggravate the genotoxicity or apoptosis.
Conclusion
Genotoxicity induced by AgNPs may be attributed to the oxidative stress induced by the nanoparticles as well as the released ions [41]. This study employed Au@Ag NR as a model to determine the distribution and release behavior of Ag after the nanoparticles enter into the cells. Considering the disparate forms of Au@Ag NR in the cell, after its exposure the Ag + was rapidly dissolved from the silver shell. Ag + and Au@Ag NR could introduce cytotoxicity and genotoxicity (clastogenicity) in the cells, and the Au@Ag NR retained in the nucleus may further release Ag + to aggravate the damage, which are mainly caused by cell cycle arrest and ROS formation (summarized in Fig. 7). Collectively, these data reveal the correlation between the intracellular accumulation, Ag + release as well as the potential genotoxicity of AgNPs.

Schematic diagram of the possible mechanism of genotoxicity introduced by AgNP in vitro
Disponibilidade de dados e materiais
All data and materials are available without restriction.
Crescimento e corrosão seletiva de estruturas multicamadas de silício / silício-germânio dopadas com fósforo para aplicação de transistores verticais
Absorção THz de banda larga de matriz de microbolômetro integrado com ressonadores de anel dividido
Nanomateriais
- A diferença entre liga de prata de tungstênio e liga de cobre de tungstênio
- Preparação de nanopartículas de mPEG-ICA carregadas com ICA e sua aplicação no tratamento de dano celular H9c2 induzido por LPS
- Examinando os papéis do tamanho da gota da emulsão e do surfactante no processo de fabricação baseado na instabilidade interfacial de nanocristais micelares
- Efeito da distribuição de nanopartículas de ouro em TiO2 nas características ópticas e elétricas de células solares sensibilizadas por corante
- A preparação da nanoestrutura de casca de gema de Au @ TiO2 e suas aplicações para degradação e detecção de azul de metileno
- Promoção do crescimento celular SH-SY5Y por nanopartículas de ouro modificadas com 6-mercaptopurina e um peptídeo penetrador de neurônio
- Profundos efeitos interfaciais em CoFe2O4 / Fe3O4 e Fe3O4 / CoFe2O4 Core / Nanopartículas de Shell
- Platycodon saponins from Platycodi Radix (Platycodon grandiflorum) para a síntese verde de nanopartículas de ouro e prata
- Nanobastões de ouro modificados com sílica conjugada com anticorpo para o diagnóstico e terapia fototérmica de Cryptococcus neoformans:um experimento in vitro
- Avaliação das propriedades antimicrobianas, apoptóticas e de entrega de genes de células cancerosas de nanopartículas de ouro protegidas por proteína sintetizadas a partir do fungo micorrízico …