


MicroRNA-133a exossômico derivado de células-tronco mesenquimais da medula óssea restringe a fibrose miocárdica e a transição epitelial-mesenquimal em ratos com miocardite viral por meio da supressão de MAML1
Resumo
Miocardite é uma doença caracterizada por inflamação localizada ou difusa do miocárdio sem tratamento eficaz. Este estudo explorou o mecanismo regulatório de microRNA-133 (miR-133) secretado por exossomo derivado de células-tronco mesenquimais da medula óssea (BMSC-Exo) na fibrose miocárdica e transição epitelial-mesenquimal (EMT) em ratos com miocardite viral (VMC) por meio da regulação como mentor 1 (MAML1). BMSCs em ratos foram isolados e cultivados para identificar seu fenótipo imunológico e capacidade osteogênica e adipogênica, e BMSC-Exo foram extraídos e identificados. Os exossomos foram obtidos por ultracentrifugação, os quais foram identificados por microscopia eletrônica de transmissão e análise de western blot. Os ratos foram injetados com o vírus Coxsackie B3 para a preparação do modelo VMC, e os cardiomiócitos foram isolados, cultivados e agrupados da mesma forma que os experimentos com animais (NC Exo , Ad-miR-133a Exo , Adas-miR-133a Exo ) Experimentos in vivo e in vitro foram conduzidos para descobrir os papéis de miR-133a exossômico e MAML1 na inflamação, apoptose, EMT, fibrose e viabilidade celular. A relação de direcionamento entre miR-133a e MAML1 foi verificada por ensaio de gene repórter de luciferase duplo. O BMSC-Exo aumentou a expressão do miR-133a em ratos VMC e melhorou efetivamente a função cardíaca do rato VMC e a fibrose miocárdica, aumentou a viabilidade dos cardiomiócitos e inibiu o processo de EMT. O miR-133a elevado em exossomos reforçou as melhorias. O miR-133a silenciado reverteu efetivamente os efeitos do BMSC-Exo em ratos VMC. miR-133a MAML1 direcionado. A inibição de MAML1 melhorou a função cardíaca e a fibrose miocárdica em ratos VMC e pode reverter o efeito de exossomos silenciados por miR-133a em ratos VMC. Nosso estudo sugere que miR-133a exossômico elevado suprime a fibrose miocárdica e a EMT em ratos com VMC por meio da regulação negativa de MAML1, inibindo assim a progressão da miocardite.
Introdução
A miocardite é considerada uma doença inflamatória das células do músculo cardíaco [1]. A miocardite é obviamente mais familiar em homens do que em mulheres [2]. A miocardite viral (VMC) é o principal fator que leva à cardiomiopatia dilatada (DCM) e morte súbita em jovens [3]. O desempenho clínico da miocardite é variado, desde estados assintomáticos com sinais e sintomas ambíguos até grave destruição do miocárdio por vírus e células imunes sofrendo de choque cardiogênico e arritmias [1]. A miocardite pode ser induzida por uma variedade de elementos infecciosos, consistindo de vírus, bactérias, Chlamydia, rickettsia, fungos e protozoários, juntamente com indutores não infecciosos. Dentre as quais, a infecção viral tem sido a causa mais comum, principalmente em crianças [4]. O vírus Coxsackie B3 (CVB3), como o vírus mais importante que leva à miocardite, pode resultar em resposta ao estresse oxidativo e apoptose na patogênese do VMC, mas o tratamento especial do VMC ainda não foi relatado [5]. Além disso, a patogênese do VMC não está bem documentada, e um tratamento clínico exato também está faltando [3]. Portanto, novos alvos são urgentemente necessários para melhorar o prognóstico da doença.
MicroRNAs (miRNAs) são RNAs endógenos não codificadores que podem regular a expressão de genes codificadores de proteínas [6]. O MiR-133a, como um dos miRNAs cardíacos específicos, está implicado no desenvolvimento do coração e em algumas doenças cardiovasculares, contendo infarto do miocárdio (IM) [7]. Além disso, miR-133a é aquele que se expressa de forma aberrante na cardiomiopatia crônica da doença de Chagas [8]. Além disso, o nível de miR-133a no miocárdio está relacionado com inflamação, função ventricular esquerda e resultado clínico na cardiomiopatia inflamatória [9]. MiRNAs foram encontrados em exossomos derivados de mastócitos de camundongo e humanos [10]. Exossomos, vesículas nanométricas liberadas pela maioria dos tipos de células são encontradas em diferentes fluidos biológicos [11]. Os exossomos podem transferir sua carga para as células receptoras, o que demonstrou alterar a composição bioquímica e as vias de sinalização das células receptoras [12, 13]. Evidências demonstraram que miRNAs exossômicos alterados estão relacionados à patogênese da miocardite induzida por CVB3 [14]. Foi abordado que miR-125b-5p exossômico de células-tronco mesenquimais da medula óssea condicionadas por hipóxia reduz a apoptose de cardiomiócitos e aumenta o reparo cardíaco isquêmico [15]. Além disso, o miR-25-3p exossômico de MSCs alivia o MI por meio da redução da apoptose dos cardiomiócitos e da resposta inflamatória [16]. De interesse, miR-133a exossômico expresso em coração está conectado à troponina-I cardíaca [17]. Mastermind-like 1 (MAML1) foi um gene a jusante de miR-133a em nosso estudo que foi relatado como envolvido em lesão de isquemia / reperfusão (I / R) miocárdica [18]. Além disso, um estudo recente mencionou que o knockdown de MAML1 possui a função antifibrótica na fibrose hepática [19].
Iluminado por estudos anteriores, é questionável se o miR-133a exossômico derivado de BMSCs poderia mediar a miocardite. Portanto, este estudo foi iniciado com a hipótese de que miR-133 transportado por exossomo derivado de BMSC (BMSC-Exo) melhora a fibrose miocárdica e a transição epitelial-mesenquimal (EMT) em ratos VMC por meio da regulação de MAML1.
Materiais e métodos
Aprovação ética
O estudo foi permitido pelo Comitê de Uso e Cuidado Institucional de Animais do Quarto Hospital Afiliado da Escola de Medicina da Universidade de Zhejiang. Os animais foram tratados com humanidade.
Isolamento de BMSCs
Os animais experimentais eram ratos machos Sprague-Dawley (SD) de grau Sprague-Dawley (SD) livres de patógenos específicos maduros (Centro de Animais Experimentais da Escola de Medicina da Universidade de Zhejiang, Zhejiang, China). Os ratos foram sacrificados por injeção intraperitoneal com pentobarbital sódico e esterilizados com álcool 75%. O fêmur e a tíbia foram retirados em uma mesa ultralimpa, os tecidos musculares e conjuntivos foram removidos e a cavidade medular foi lavada repetidamente com Meio Eagle Modificado por Dulbecco (DMEM) com baixo teor de glicose. O líquido foi centrifugado para coletar os precipitados que foram ressuspensos e incubados por 24 h (o meio foi trocado a cada 2-3 dias). Ao crescer para a fase logarítmica, as BMSCs foram separadas com 0,25% de tripsina (Gibco, Carlsbad, Califórnia, EUA), centrifugadas e ressuspensas em solução de cultura MSC (Cyagen Biosciences Inc., Guangzhou, China). A suspensão foi passada na proporção de 1:2. A operação acima foi repetida, e as BMSCs da 4ª passagem foram usadas para experiências subsequentes.
Identificação de BMSCs
Os antígenos de superfície das BMSCs de 4ª passagem em crescimento logarítmico foram identificados por citometria de fluxo. As BMSCs foram separadas com 0,25% de tripsina (1 mL) contendo ácido etilenodiaminotetracético, centrifugadas, ressuspensas com solução salina tamponada com fosfato (PBS) e centrifugadas a 151 g. As BMSCs foram então ressuspensas com PBS contendo 2% de soro fetal bovino fresco (FBS) (Gibco) para fazer uma suspensão de célula única. FITC-CD34, PE-CD29 e anticorpo monoclonal PE-CD44 (5 μL cada, BD Biosciences, Franklin Lakes, NJ, EUA) foram incubados com a suspensão de células (100 μL), centrifugados a 151 g, ressuspensos com 500 μL de PBS contendo 1% de paraformaldeído e fixado por 30 min. Os marcadores de fundo foram identificados por meio do uso de anticorpos monoclonais do controle de homótipo.
Citometria de fluxo:a suspensão unicelular foi fixada e centrifugada a 151 g. Em seguida, as BMSCs foram ressuspensas com PBS contendo 1% de paraformaldeído, testadas no citômetro de fluxo MACS Quart e analisadas pelo software correspondente.
Indução de Osteogênese e Adipogênese de BMSCs
As BMSCs na 4ª passagem foram semeadas em placas de 6 poços com 200 células / mL. A solução de indução de osteoblastos e a solução de indução adipogênica (Cyagen Biosciences Inc.) foram adicionadas a BMSCs de confluência de 60–70%. BMSCs em outros dois poços não foram adicionados com líquidos de indução como controles. As BMSCs foram induzidas por 14 dias e fixadas com paraformaldeído 4%. Em seguida, os osteoblastos e adipócitos diferenciados foram implementados com coloração com vermelho de Alizarina e coloração com vermelho de óleo O (Wuhan Pulande Biological Technology Co., Ltd., Wuhan, China) e observados ao microscópio.
Isolamento e identificação de exossomos
As BMSCs na 4ª passagem foram cultivadas por 48 h para colher o sobrenadante que foi então centrifugado (800 ge 2.000 g), filtrado com membranas de filtro de 0,22 μm e 100.000 MW e centrifugado (100.000 g) para coletar os precipitados. Em seguida, os precipitados foram ressuspensos com PBS, centrifugados a 100,00 g novamente para obter precipitação do exossomo. A suspensão BMSC-Exo em PBS foi submetida à detecção de concentração por ácido bicinconínico (BCA) e detecção de proteína criadora de exossomo (CD63, CD81 e CD9) por análise de western blot (Proteintech, Chicago, IL, EUA).
A infecção por adenovírus recombinante medeia a modificação do gene miR-133a de BMSCs
BMSCs foram passados durante a noite. O controle normal (quantidade igual de PBS), o controle negativo miR-133a (NC), a superexpressão de miR-133a (Ad-miR-133a) e miR-133a de baixa expressão (Adas-miR-133a) ( Hanbio Biotechnology Co., Ltd., Shanghai, China) foram transfectados com BMSCs em linha com 100 multiplicidade de infecção (MOI). Os BMSCs foram cultivados e os exossomos correspondentes (NC Exo , NC Exo , Ad-miR-133a Exo e Adas-miR-133a Exo ) foram obtidos por ultracentrifugação [20].
Estabelecimento do modelo VMC em ratos e agrupamento de animais experimentais
Ratos SD machos maduros de grau SPF foram divididos em 10 grupos, com oito ratos cada. O Coxsackievirus B3 (CVB3) foi fornecido pelo Instituto de Biotecnologia Médica da Academia Chinesa de Ciências Médicas (Pequim, China).
CVB3 (10 mg / kg) foi injetado intraperitonealmente em ratos enquanto PBS ou BMSC-Exo (100 μg) foi injetado através da veia da cauda. Ratos normais para controles foram injetados com solução de cultura CVB3 e PBS. Ratos injetados com 10 mg / kg de CVB3 foram ainda injetados com PBS, MSC exo , NC Exo , Adas-miR-133a Exo , Ad-miR-133a Exo , si-NC ou si-MAML1 (RIBOBIO, Guangzhou, China).
Os ratos foram injetados continuamente por 7 dias e o sangue do globo ocular foi obtido. O sangue foi centrifugado e o soro coletado, sub-embalado e armazenado a -20 ° C. Após a eutanásia dos ratos, as amostras do coração foram retiradas, fixadas com formol a 10%, desidratadas com álcool gradiente, depuradas com xilol e incluídas em parafina, seccionadas para observação histológica. Uma parte das seções foram colocadas a -80 ° C como os materiais dos experimentos de biologia molecular.
Ecocardiografia
No 7º dia após a injeção do vírus, os ratos foram injetados intraperitonealmente com pentobarbital sódico 25 mg / kg. Após a anestesia completa, a derivação do membro da máquina de eletrocardiograma conectada à agulha do eletrodo foi inserida subcutaneamente nas extremidades dos membros dos ratos, e o eletrocardiograma da derivação do membro foi registrado. Em seguida, os ratos foram fixados em decúbito dorsal ligeiramente à esquerda, o tórax foi depilado e o eletrocardiograma de derivação II foi conectado para obtenção do espectro Doppler do pulso de fluxo sanguíneo da aorta no corte paraesternal de quatro câmaras. Os indicadores incluíram a espessura da parede posterior do ventrículo esquerdo (LVPW), o diâmetro sistólico final do ventrículo esquerdo (LVIDs), a fração de encurtamento do ventrículo esquerdo (FS) e a fração de ejeção do ventrículo esquerdo (LVEF).
Coloração com Hematoxilina-Eosina (HE)
Os tecidos foram fixados com paraformaldeído 4%, desidratados, clarificados e incluídos em parafina. Em seguida, as seções de 4 μm foram desceradas, coradas com hematoxilina (Servicebio, Wuhan, China), diferenciadas com álcool ácido clorídrico a 1%, retornadas ao azul e coradas com eosina, desidratadas, depuradas com xileno, seladas com goma neutra, e observada sob um microscópio óptico (Olympus, Tóquio, Japão).
Coloração com Colágeno Masson
Os cortes de parafina foram descerados, corados com hematoxilina por menos de 2 min, corados com solução magenta de Lichun e rapidamente enxaguados com solução de ácido acético glacial 0,5%. Em seguida, as seções foram coradas com solução aquosa de fosfato de alumínio a 1%, coradas de vermelho escuro a vermelho vivo a rosa e observadas ao microscópio. Em seguida, os cortes foram corados com azul de anilina (Pulande), convencionalmente desidratado com xilol e lacrados. O software de análise de imagens médicas Image-Proplus 6.0 foi usado para medir a área de coloração positiva das fibras colágenas e a fração de volume de colágeno (CVF) =área colágena / área total do campo. O local da coloração e a cor das fibras colágenas foram diferenciadas (os cardiomiócitos eram vermelhos e as fibras colágenas eram com faixas azuis ou estruturas homogêneas no espaço intercelular).
Coloração terminal de desoxinucleotidil transferase mediada por desoxiuridina trifosfato-biotina Nick (TUNEL) coloração final
As seções de parafina foram descascadas, colocadas no tampão de citrato e cozidas a 350 W por 10 min. As seções foram adicionadas com 50 μL de solução TUNEL, unidas com 50 μL de agente de conversão-peroxidase, desenvolvido com DAB e observadas ao microscópio. Os cortes foram colocados em hematoxilina, embebidos em etanol I – II a 95%, unidos com etanol anidro I – II, xileno I – II e selados. Os resultados foram analisados ao microscópio óptico.
Ensaio de imunoabsorção enzimática (ELISA)
Fator de necrose tumoral α (TNF-α), interleucina (IL) -1β e IL-6 foram detectados por kits de ELISA (BOSTER Biological Technology Co. Ltd., Wuhan, China). O sangue do globo ocular foi centrifugado a 604 g para coletar o soro superior. O sobrenadante obtido por centrifugação do meio de cultura de células foi detectado no experimento de células. Havia sete gradientes de concentração no padrão de diluição das amostras. O poço em branco foi unido ao diluente de amostra e outro poço foi adicionado com tetrametilbenzidina (TMB), dois poços duplicados foram ajustados para cada concentração. Os poços das amostras foram unidos com 50 μL de diluente de amostra e as amostras por sua vez. Cada poço reagiu com 100 μL de anticorpo primário (exceto poço TMB) por 1 h, bem como com 300 μL de solução salina tamponada com tris 0,01 M (TBS) e 100 μL de solução de trabalho do Complexo Avidina-Biotina-Peroxidase (exceto poço TMB ) Em seguida, cada poço foi adicionado com 300 μL de TBS 0,01 M e incubado com 100 μL de TMB. O valor da densidade óptica (DO) e a concentração de cada poço foram medidos imediatamente e a curva padrão foi desenhada.
Reação em cadeia da polimerase quantitativa com transcrição reversa (RT-qPCR)
MiR-133a, colágeno Ι, colágeno III, α-SMA, TGF-β1, CTGF, E-caderina e expressão de FSP-1 em tecidos miocárdicos e cardiomiócitos foram detectados por RT-qPCR. O RNA total foi extraído de cardiomiócitos ou tecidos do miocárdio e reversamente transcrito para cDNA via kit de extração de RNA (Takara, Dalian, China), e os primers de RT-PCR foram sintetizados via Invitrogen (Guangzhou, China), as sequências são mostradas na Tabela 1. A expressão quantitativa relativa do gene foi analisada usando gliceraldeído-3-fosfato desidrogenase (GAPDH) ou U6 como os genes de controle de carregamento de acordo com 2 - △△ Ct método.
Análise de Western Blot
Os ratos foram sacrificados com anestesia. Os tecidos miocárdicos foram congelados e moídos em nitrogênio líquido. Em seguida, a solução estoque de fenilmetanossulfonilfluoreto de inibidores de protease foi misturada com tampão de lise celular na proporção de 1:100 (Beyotime Biotechnology Co., Ltd., Shanghai, China). As amostras foram lisadas com solução mista e as proteínas das células foram extraídas. A concentração total de proteína foi detectada com o kit BCA. As amostras foram misturadas com tampão de carregamento 5 × a 4:1, implementado com banho-maria em ebulição por 10 min, banhado em gelo e centrifugado. Foi realizada a separação por eletroforese e as proteínas foram transferidas para membrana de fluoreto de polivinilideno (Servicebio) com solução de transferência elétrica. Em seguida, a membrana foi bloqueada com leite em pó desnatado a 5% e unida aos anticorpos primários CD63, CD81 e CD9 (anticorpos policlonais de coelho anti-rato da Proteintech, 1:100), MAML1 (ab65090, Abcam, MA, USA, 1:1000) e GAPDH (Santa Cruz Biotechnology, Inc, Santa Cruz, CA, EUA, 1:1000). Em seguida, a membrana foi gotejada com o anticorpo secundário, IgG marcada com peroxidase de rábano (Cell Signaling Technology, Beverly, MA, EUA, 1:1000) e imersa em solução de reação de quimioluminescência aprimorada (Pierce, Rockford, IL, EUA). Com GAPDH como controle de carregamento, as imagens de impressão de proteína foram analisadas com o software ImageJ2x.
Cultura e passagem de cardiomiócitos
Ratos SD com 3–5 dias de idade (Centro de Animais Experimentais da Escola de Medicina da Universidade de Zhejiang, Zhejiang, China) foram coletados. A parte ventricular foi enxaguada com solução salina balanceada de Hank pré-resfriada, cortada em pequenos pedaços e separada com 0,25% de tripsina. As peças foram adicionadas com uma quantidade adequada de meio completo a 10% para término da separação e centrifugadas a 151 g. DMEM contendo 20% de FBS foi aplicado para ressuspensão celular. Os cardiomiócitos foram purificados por método diferencial de aderência, e a taxa de sobrevivência foi observada por meio de coloração com azul de tripano, os cardiomiócitos sobreviventes foram cultivados. Após 24 horas, os cardiomiócitos aderiram à parede e começaram a pulsar. Após 72 h, os pseudópodes expandiram.
Construção do modelo VMC de cardiomiócito
Cardiomiócitos na 4ª passagem na fase de crescimento logarítmico foram selecionados e infectados com MSC Exo , NC Exo , Adas-miR-133a Exo e Ad-miR-133a Exo . A solução de vírus 100 Tcid50 CVB3 (100 μL) foi adicionada às células para induzir um modelo de célula VMC. Ao mesmo tempo, quantidade igual de solução de manutenção foi adicionada às células para um controle, e os exossomos correspondentes foram unidos aos cardiomiócitos após infecção de 1 hora para cultura de 47 horas.
Ensaio de kit de contagem de células (CCK) -8
O kit de detecção de células CCK-8 (Beyotime) foi aplicado para detectar a taxa de sobrevivência dos cardiomiócitos. Ao crescer para a fase logarítmica, as células foram destacadas com 0,25% de tripsina e semeadas em placas de cultura de células de 96 poços a 2,5 × 10 4 células / por poço. Juntamente com a solução de CCK-8 (10 μL / poço), as células foram cultivadas continuamente por 1–4 h, e a DO 450 nm o valor foi medido através de um leitor de microplaca.
Citometria de fluxo
O método de dupla coloração com anexinV-APC / iodeto de propídio (PI) foi aplicado para detectar a apoptose celular. As células foram centrifugadas, ressuspensas com 250 μL de tampão de ligação (4 mL de tampão de ligação + 12 mL de água desionizada) e ajustadas para 1 × 10 6 células / mL. A suspensão de células de 100 μL foi adicionada com 5 μL de Anexina V-APC (BD Biosciences) e 5 μL de solução de PI (BD Biosciences), carregada no citômetro de fluxo e analisada automaticamente por um computador.
Ensaio do gene Dual Luciferase Reporter
A sequência de tipo selvagem (wt) ou tipo mutante (mut) da região 3-não traduzida de MAML1 (UTR) foi clonada no vetor pGL3-M (Promega, WI, EUA), depois MAML1-3-UTR-wt ou MAML1- 3-UTR-mut foram gerados. Os vetores, juntamente com miR-133a mimetizador ou NC, foram co-transfectados em cardiomiócitos via Lipofectamine 2000. A atividade da luciferase foi testada 48 h mais tarde pelo sistema de gene repórter de luciferase dual (Promega) [21]
Ensaio de imunoprecipitação de RNA (RIP)
O kit RIP (Millipore, EUA) foi usado para detectar a ligação de MAML1 e miR-133a. As células foram lisadas por tampão de ensaio de radioimunoprecipitação (P0013B, Beyotime, Shanghai, China), centrifugadas a 1400 ge incubadas com anticorpos para co-precipitar. Grânulos magnéticos (50 μL) foram ressuspensos em 100 μL de tampão de lavagem RIP e incubados com 5 g de anticorpo anti-MAML1 (1 g / mL, ab155786) ou IgG (1:100, ab172730). O complexo magnético esfera-anticorpo foi ressuspenso em 900 μL de tampão de lavagem RIP, interagiu com 100 μL de extrato celular, digerido com proteinase K e detectado por RT-qPCR [22].
Análise estatística
O software estatístico SPSS 21.0 (IBM Corp. Armonk, NY, EUA) foi aplicado para análise dos dados. Os dados de medição foram expressos como média ± desvio padrão. O teste t foi aplicado para comparações entre dois grupos. A análise de variância (ANOVA) unilateral foi usada para comparação entre os grupos e o teste post hoc de Tukey para comparações entre pares. Os preditores eram mantidos se fossem significativos em um P valor de 0,05 ou menor.
Resultados
Identificação de BMSCs e BMSC-Exo
Microscopicamente, as BMSCs eram fusiformes e arredondadas, e aderidas à parede em um vórtice ou padrão radial (Fig. 1A). Após a coloração com óleo vermelho O de indução de adipogênese, as gotículas lipídicas da 4ª BMSCs eram vermelhas e havia gotículas lipídicas redondas de tamanhos diferentes (Fig. 1B). Após a indução da osteogênese, as células que expressavam nódulos calcificados mostraram-se vermelhas após a coloração com vermelho de alizarina e distribuição irregular de nódulos calcificados e células sobrepostas (fig. 1C). A citometria de fluxo manifestou que os marcadores de MSC CD29 e CD44 (> 95%) foram expressos, mas o antígeno de superfície de células-tronco hematopoiéticas CD34 (<95%) não foi expresso (Fig. 1D). Estes resultados manifestaram que os BMSCs eram de alta pureza e estavam em conformidade com os padrões MSC da International Society of Cell Therapy.
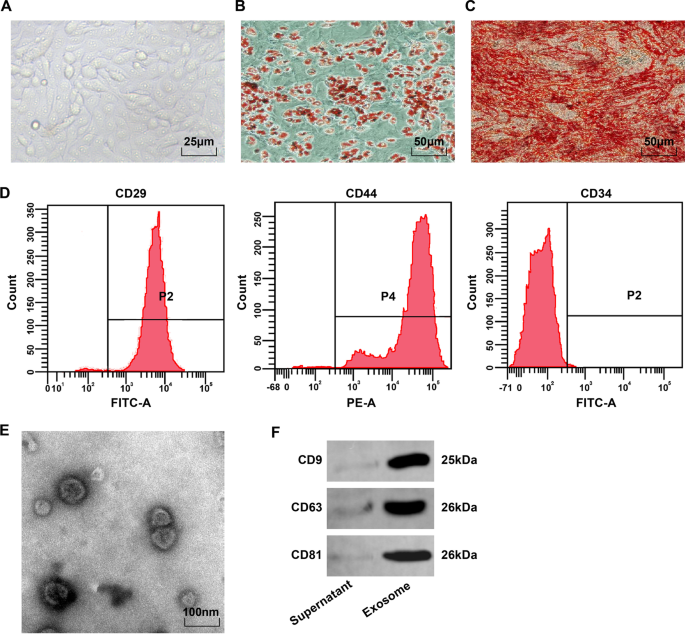
Observação do fenótipo BMSC e identificação BMSC-Exo. A Observação morfológica de BMSCs na 4ª passagem; B Os resultados da coloração com óleo vermelho O de adipócitos; C Os resultados da coloração com vermelho de alizarina de osteoblastos; D O fenótipo de BMSC detectado por meio de citometria de fluxo. E Observação por microscópio eletrônico de BMSC-Exo; F Bandas de proteína de CD9, CD63 e CD81
O microscópio eletrônico de transmissão observou que BMSC-Exos eram vesículas ovais com estrutura membranosa periférica clara, tamanhos diferentes e diâmetro de 40–100 nm (Fig. 1E). A análise de Western blot demonstrou que os produtos extraídos expressaram exossomos originados de proteínas características de CD9, CD63 e CD81 (Fig. 1F).
O miR-133a exossômico elevado melhora os sintomas de miocardite
A transfecção de BMSCs com miR-133a adenovírus recombinante foi observada (Fig. 2A). Um grande número de expressão de fluorescência verde de NC, Ad-miR-133a e Adas-miR-133a foi observada sob o microscópio de fluorescência invertido, indicando que o vetor de adenovírus recombinante poderia efetivamente transfectar BMSCs. Para testar a eficiência de transfecção de miR-133a, a expressão de miR-133a em BMSCs e seus exossomos foi medida por RT-qPCR. Foi descoberto que a regulação positiva de miR-133a aumentou a expressão de miR-133a, enquanto a regulação negativa de miR-133a diminuiu a expressão de miR-133a (Fig. 2B). Posteriormente, injetamos exossomos contendo miR-133a em ratos. Sob o microscópio de fluorescência invertido, a expressão de fluorescência verde foi observada em ratos VMC após o tratamento de NC Exo , Ad-miR-133a Exo ou Adas-miR-133a Exo , indicando que o vetor de adenovírus recombinante infectou tecidos do miocárdio de ratos (Fig. 2C). O experimento RT-qPCR também descobriu que a expressão de miR-133a em ratos VMC foi aparentemente diminuída; A expressão do miR-133a foi obviamente elevada em ratos VMC injetados com Ad-miR-133a Exo mas diminuiu em ratos VMC injetados com Adas-miR-133a Exo (Fig. 2D). Quanto às condições gerais dos ratos, observou-se que as condições gerais dos ratos controle normais eram normais, e as características de VMC foram aparentemente expressas em ratos VMC e ratos VMC injetados com Adas-miR-133a Exo , como cabelo áspero e desordenado, dispneia e pouca dieta. Em ratos VMC tratados com MSC Exo , NC Exo e Ad-miR-133a Exo , esses sinais foram melhorados em graus diferentes. O peso dos ratos VMC diminuiu continuamente a partir de 1 dia após a infecção e injeção de MSC Exo , NC Exo ou Ad-miR-133a Exo aumentou o peso dos ratos. O peso dos ratos VMC tratados com Ad-miR-133a Exo foi aumentado obviamente e o peso dos ratos VMC injetados com Adas-miR-133a Exo foi claramente diminuída (Fig. 2E).
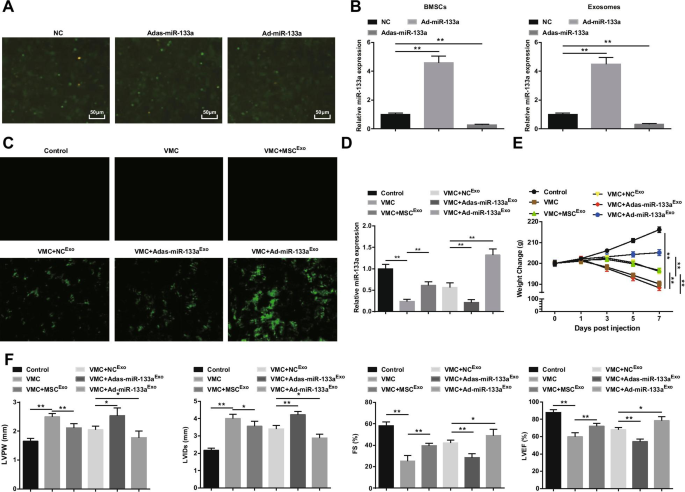
O miR-133a exossômico regulado positivamente alivia a miocardite. A Transfecção de BMSCs de adenovírus recombinante miR-133a; B Detecção de RT-qPCR da expressão de miR-133a em BMSCs e seus exossomos após regular miR-133a; C eficiência de transfecção miR-133a testada através de microscópio de fluorescência invertido; D A expressão relativa de miR-133a em tecidos miocárdicos testados via RT-qPCR; E Mudança de peso dos ratos em cada grupo; F Determinação de LVPW, LVIDs, FS e LVEF em ratos de cada grupo. * P <0,05; ** P <0,001
A observação da função miocárdica sugeriu que (fig. 2F) os ratos VMC apresentaram aumento do LVPW e LVIDs e diminuição do FS e LVEF. Após a injeção de exossomo, LVPW e LVIDs declinaram e, obviamente, FS e LVEF elevados mostraram-se em ratos VMC. Adas-miR-133a Exo tratamento prejudicado enquanto Ad-miR-133a Exo melhora da função miocárdica em ratos VMC.
O miR-133a exossômico regulado positivamente inibe a inflamação nos tecidos miocárdicos de ratos VMC
A coloração HE manifestou que as fibras miocárdicas em ratos de controle normais estavam estreitamente organizadas e não havia infiltração de células inflamatórias no mesênquima. Os cardiomiócitos nos ratos VMC estavam desorganizados e o mesênquima infiltrado por grande número de células inflamatórias. Os cardiomiócitos em ratos VMC injetados com MSC Exo ou NC Exo estavam organizados em ordem, com uma pequena quantidade de células inflamatórias infiltrando-se no mesênquima. Os cardiomiócitos em ratos VMC após Adas-miR-133a Exo o tratamento estava organizado de maneira desordenada e as células inflamatórias do mesênquima estavam infiltradas. Os cardiomiócitos em ratos VMC tratados com Ad-miR-133a Exo foram organizados em ordem sem infiltração de células inflamatórias óbvias (Fig. 3A).
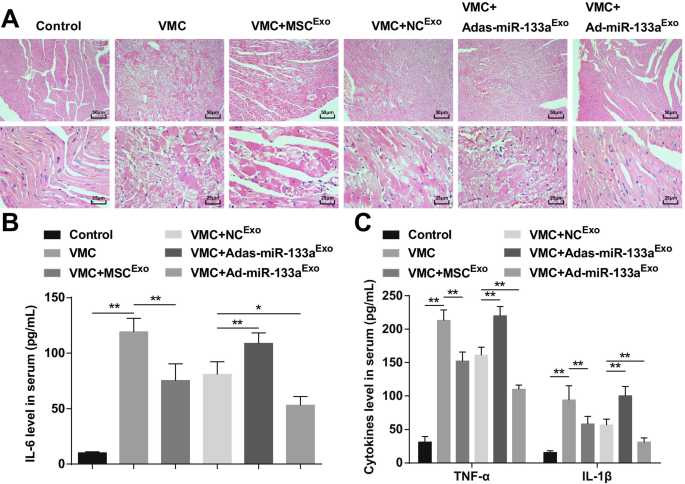
O aumento do miR-133a exossômico restringe a inflamação nos tecidos do miocárdio com VMC. A Coloração HE do tecido do miocárdio de rato em cada grupo; B A expressão de IL-6 no soro testada via ELISA; C Expressão de TNF-α e IL-1β no soro testada via ELISA. * P <0,05; ** P <0,001
ELISA indicou que (Fig. 3B, C) fatores inflamatórios (TNF-α, IL-1β e IL-6) estavam obviamente aumentados em ratos VMC. Os ratos VMC injetados com Ad-miR-133a Exo tinham níveis reduzidos de fatores inflamatórios. Adas-miR-133a Exo o tratamento causou fatores inflamatórios elevados em ratos VMC.
MiR-133a exossômico elevado diminui a CVF em tecidos miocárdicos de ratos com VMC
A coloração de Masson revelou que as fibras miocárdicas em ratos normais estavam organizadas de maneira próxima, quase sem fibras colágenas azuis. Após a injeção de CVB3, os cardiomiócitos estavam hipertróficos, com hiperplasia do tecido conjuntivo e um grande número de fibras colágenas azuis, e o CVF estava obviamente elevado. Tratados com exossomos, os cardiomiócitos foram organizados em ordem, a hiperplasia do tecido conjuntivo intercelular diminuiu, as fibras de colágeno azul e a CVF estavam claramente diminuídas. O espaço intercelular do miocárdio de ratos VMC injetados com Adas-miR-133a Exo foi alargado, as células estavam obviamente aumentadas, as fibras de colágeno azul e CVF estavam claramente aumentadas; o espaço intercelular foi diminuído, a distribuição de fibras de colágeno azuis e o CVF foram diminuídos de ratos VMC com Ad-miR-133a Exo tratamento (Fig. 4A, B).
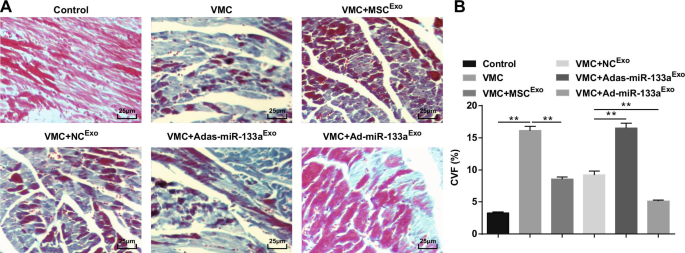
O miR-133a exossômico regulado positivamente diminui a CVF em tecidos do miocárdio de ratos com VMC. A Coloração de Masson de tecidos miocárdicos em ratos; B Fração de volume de colágeno de ratos em cada grupo. ** P <0,001
MiR-133a exossômico aumentado reduz a expressão de colágeno I, colágeno III, TGF-β1 e CTGF em tecidos miocárdicos de ratos com VMC
O colágeno I e o colágeno III são os principais componentes do colágeno, que se distribuem principalmente nas junções e membranas celulares, na substância intercelular e no citoplasma. TGF-β1 e CTGF são as proteínas marcantes da fibrose. Os achados de RT-qPCR demonstraram que os níveis de expressão de colágeno I, colágeno III, TGF-β1 e CTGF mRNA aumentaram em ratos VMC, mas diminuíram após o tratamento com exossomo. Os ratos VMC tratados com Ad-miR-133a Exo diminuíram os níveis de expressão de mRNA de colágeno I, colágeno III, TGF-β1 e CTGF enquanto os ratos VMC após Adas-miR-133a Exo o tratamento mostrou a situação oposta (Fig. 5A-C).
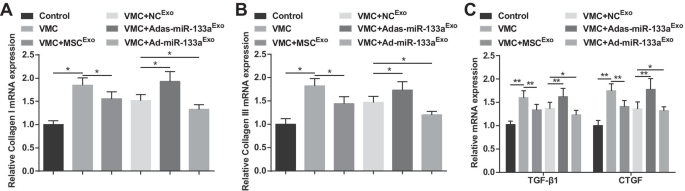
O miR-133a exossômico elevado reduz a expressão de mRNA de colágeno I, colágeno III, TGF-β1 e CTGF em tecidos miocárdicos de ratos com VMC. A A expressão do mRNA do colágeno I em tecidos miocárdicos de ratos foi detectada por RT-qPCR; B A expressão do mRNA do colágeno III em tecidos miocárdicos de ratos foi detectada por RT-qPCR; C A expressão de mRNA de TGF-β1 e CTGF em tecidos miocárdicos de ratos foi detectada por RT-qPCR. * P <0,05; ** P <0,001
O miR-133a exossômico regulado positivamente inibe a apoptose de cardiomiócitos em tecidos miocárdicos de ratos com VMC
A coloração de TUNEL mostrou que os cardiomiócitos apoptóticos eram preto acastanhado ou amarelo acastanhado com condensação nuclear. O número de células apoptóticas foi aumentado em ratos VMC, o que seria atenuado pelo tratamento com exossomo. Os ratos VMC injetados com Ad-miR-133a Exo teve número reduzido de células apoptóticas e aquelas injetadas com Adas-miR-133a Exo tinha aumento do número de células apoptóticas (Fig. 6A, B).
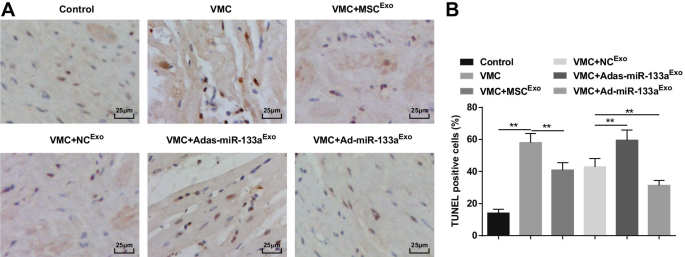
O aumento do miR-133a exossômico inibe a apoptose de cardiomiócitos em tecidos miocárdicos de ratos com VMC. A Coloração TUNEL de tecidos do miocárdio de rato em cada grupo; B O número de células TUNEL positivas em cada grupo. ** P <0,001
O miR-133a exossômico elevado diminui a EMT em tecidos miocárdicos de ratos com VMC
E-caderina, α-SMA e FSP-1 são indicadores-chave de EMT. Os resultados de RT-qPCR demonstraram que os níveis de expressão de mRNA de α-SMA e FSP-1 estavam elevados e o nível de expressão de mRNA da caderina E estava diminuído em ratos VMC. Além disso, os níveis de expressão de mRNA de α-SMA e FSP-1 foram reduzidos e o nível de expressão de mRNA da caderina E foi aumentado em ratos VMC após o tratamento com exossomo. Os níveis de expressão de mRNA de α-SMA e FSP-1 foram elevados e o nível de expressão de mRNA de E-caderina diminuiu em ratos VMC tratados com Adas-miR-133a Exo , enquanto a expressão desses indicadores foi oposta em ratos VMC injetados com Ad-miR-133a Exo (Fig. 7A, B).
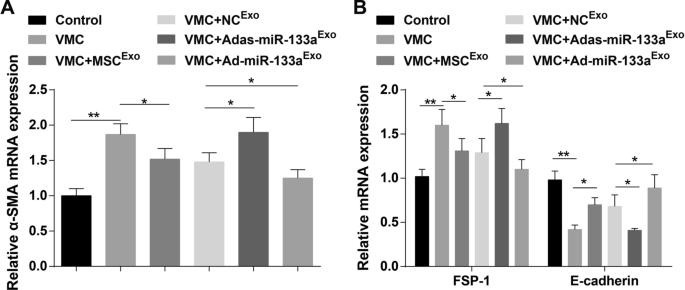
O miR-133a exossômico regulado positivamente reprime a EMT nos tecidos do miocárdio de ratos com VMC. A A expressão de mRNA de α-SMA em tecidos do miocárdio de rato em cada grupo foi detectada por RT-qPCR; B A expressão de mRNA de FSP-1 e E-caderina em tecidos do miocárdio de rato em cada grupo foi detectada por RT-qPCR. * P <0,05; ** P <0,001
MiR-133a exossômico regulado para cima diminui a inflamação dos cardiomiócitos em VMC
Como resultado, a microscopia de fluorescência capturou a expressão fluorescente verde em ratos VMC tratados com NC Exo , Ad-miR-133a Exo ou Adas-miR-133a Exo , indicando que o vetor de adenovírus recombinante infectou cardiomiócitos de ratos (Fig. 8A). RT-qPCR e ELISA descobriram que (Fig. 8B, D) a expressão de miR-133a foi reduzida e fatores inflamatórios (TNF-α, IL-1β e IL-6) foram aumentados em ratos VMC, que seriam revertidos por tratamento com exossomo . Os ratos VMC tratados com Ad-miR-133a Exo tinham miR-133a regulado positivamente e fatores inflamatórios diminuídos em ratos VMC, enquanto aqueles tratados com Adas-miR-133a Exo apresentou diminuição do miR-133a e aumento dos níveis de fatores inflamatórios em ratos VMC.
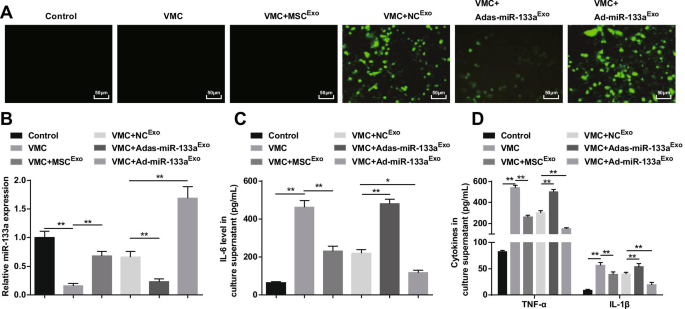
O miR-133a exossômico elevado restringe a inflamação dos cardiomiócitos em VMC. A eficiência de transfecção miR-133a testada através de microscópio de fluorescência invertido; B A expressão relativa de miR-133a em cardiomiócitos de ratos em cada grupo; C Expressão de IL-6 em sobrenadante de cultura de cardiomiócitos em cada grupo; D Expressão de TNF-α e IL-1β no sobrenadante de cultura de cardiomiócitos em cada grupo. * P <0,05; ** P <0,001
O miR-133a exossômico elevado promove a viabilidade celular e reprime a apoptose de cardiomiócitos em VMC
A apoptose e a viabilidade celular foram detectadas via coloração dupla com AnnexinV-APC / PI e ensaio CCK-8. Os resultados revelaram que houve um aumento óbvio na taxa de apoptose, uma diminuição na viabilidade celular dos cardiomiócitos em ratos VMC. O tratamento com exossomo reduziu a taxa de apoptose e aumentou a viabilidade dos cardiomiócitos. Adas-miR-133a Exo aumentou a taxa de apoptose e interrompeu a viabilidade dos cardiomiócitos em ratos VMC. Ad-miR-133a Exo o tratamento funcionou com efeitos opostos nos cardiomiócitos de ratos VMC (Fig. 9A-C).
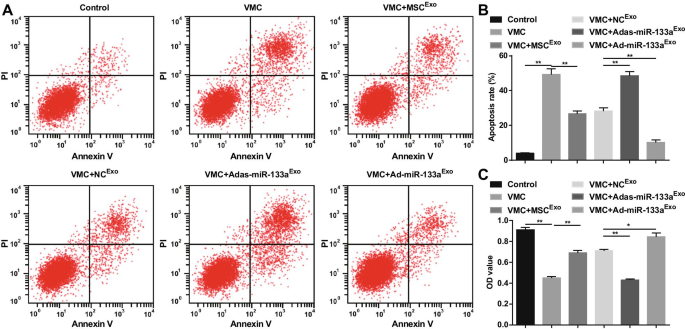
O aumento do miR-133a exossômico promove a viabilidade e reprime a apoptose em cardiomiócitos em VMC. A A apoptose dos cardiomiócitos detectada por citometria de fluxo; B Resultados de quantificação de A; C A viabilidade celular detectada através do ensaio CCK-8. * P <0,05; ** P <0,001
miR-133a Alvos MAML1
Foi relatado que o miRNA-193b regulado positivamente reduz o dano de I / R do miocárdio ao direcionar o MAML1 [18]. Com base nisso, fizemos a triagem cruzada de genes downstream de miR-133a por meio de sites de bioinformática PITA, miRanda, PicTar, microT e miRmap, e selecionamos MAML1 como um alvo de miR-133a (Fig. 10A). Construímos MAML1-wt ou MAML1-mut e cardiomiócitos co-transfectados com miR-133a mimetizador ou NC. Os resultados mostraram que o miR-133a mimetizador reduziu a atividade da luciferase de MAML1-wt (Fig. 10B). O experimento RIP verificou ainda a relação de direcionamento entre miR-133a e MAML1 (Fig. 10C). A detecção de RT-qPCR e Western blot da expressão de MAML1 mostrou que a expressão de MAML1 foi diminuída em cardiomiócitos transfectados com miR-133a mimetizador (Fig. 10D, E).
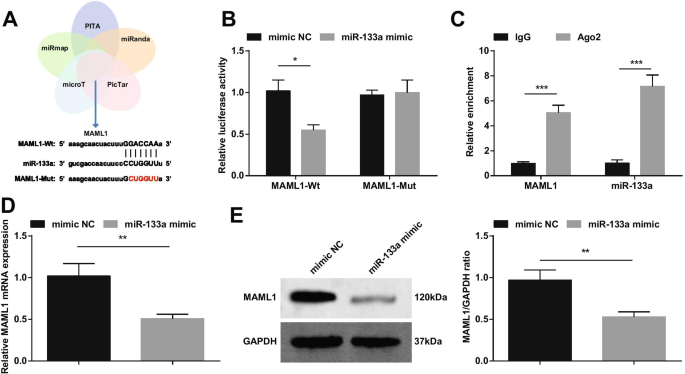
miR-133a tem como alvo MAML1. A alvos do miR-133a previstos em sites de bioinformática; B A relação de direcionamento entre miR-133a e MAML1 verificada por experimento de gene repórter de luciferase dupla; C A relação de direcionamento entre miR-133a e MAML1 verificada pelo experimento RIP; D / E A expressão de MAML1 muda após a regulação positiva de miR-133a detectada por RT-qPCR e Western blot. * P <0,05; ** P <0,01; *** P <0,001
A inibição de MAML1 tem um efeito protetor em ratos com miocardite e reverte o efeito de exossomos inibidos por miR-133a em ratos com VMC
Para estudar ainda mais o efeito de MAML1 regulado por miR-133a em ratos com VMC, injetamos adenovírus si-MAML1 ou si-NC em ratos VMC ou ratos VMC que foram tratados com exossomos silenciados por miR-133a. O sucesso da injeção foi validado por RT-qPCR e Western blot (Fig. 11A, B). Os resultados manifestaram que a injeção de si-MAML1, o peso dos ratos VMC foi aumentado (Fig. 11C), a função cardíaca foi melhorada (Fig. 11D-G), a patologia do tecido miocárdico e a fibrose foram atenuadas (Fig. 12A-C), inflamação sérica (Fig. 12D, E) e apoptose de cardiomiócitos (Fig. 13A-G) foram inibidas. Além disso, os efeitos deletérios dos exossomos silenciados com miR-133a em ratos VMC foram revertidos após a injeção de si-MAML1.
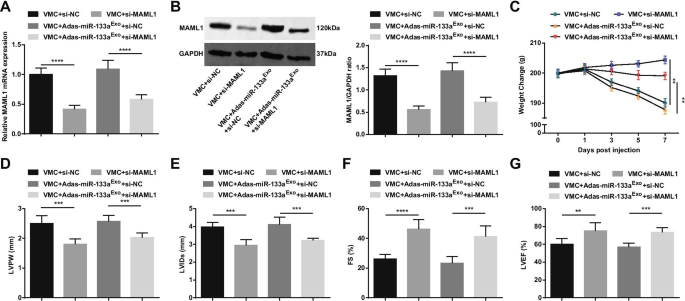
A inibição de MAML1 tem um efeito protetor em ratos com miocardite e pode reverter o efeito de exossomos silenciados por miR-133a em ratos com VMC. A / B Expressão de MAML1 em tecido miocárdico de ratos detectada por RT-qPCR e Western blot; C. Mudança de peso de ratos; D - G Determinação de LVPW, LVIDs, FS e LVEF em ratos; ** P <0,01; *** P <0,001; **** P <0,0001
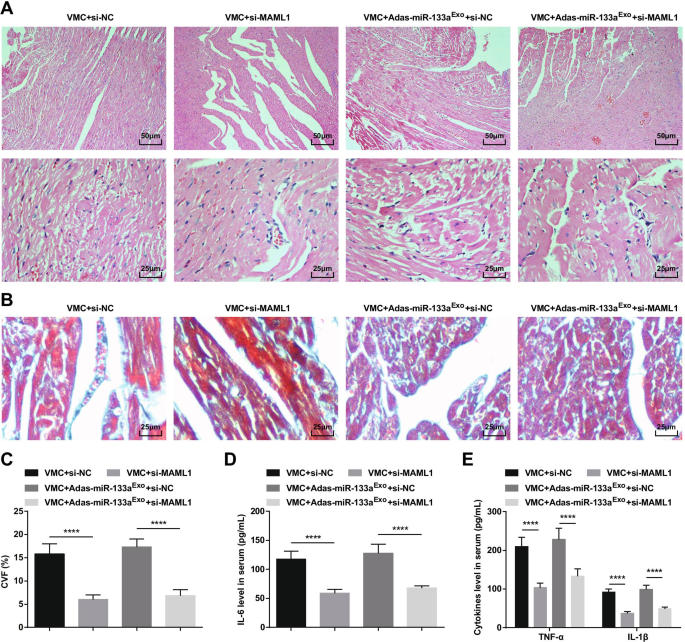
A inibição de MAML1 pode reverter o efeito de exossomos silenciados por miR-133a em ratos com VMC. A Coloração HE de tecido do miocárdio de rato; B Coloração de Masson de tecidos miocárdicos em ratos; C CVF de ratos; D A expressão de IL-6 no soro testada via ELISA; E A expressão de TNF-α e IL-1β no soro testada via ELISA. **** P <0,0001
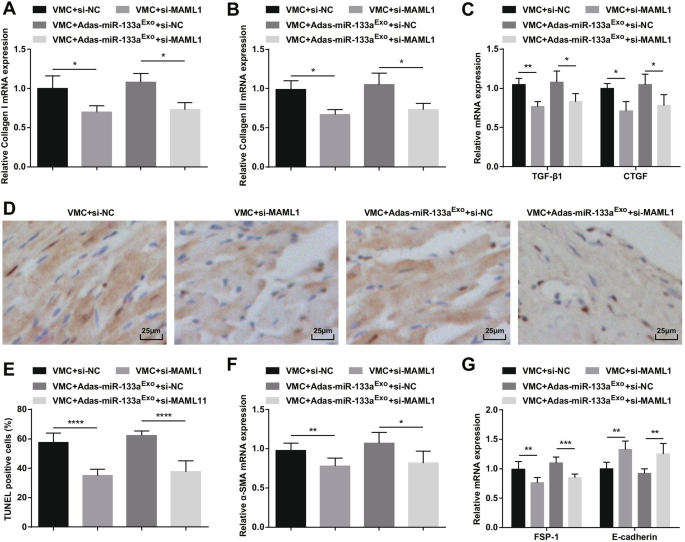
A inibição de MAML1 pode reverter o efeito dos exossomos inibidores de miR-133a em ratos com VMC. A A expressão do mRNA do colágeno I em tecidos miocárdicos de ratos foi detectada por RT-qPCR; B A expressão do mRNA do colágeno III em tecidos miocárdicos de ratos foi detectada por RT-qPCR; C A expressão de mRNA de TGF-β1 e CTGF em tecidos miocárdicos de ratos foi detectada por RT-qPCR; D Coloração TUNEL de tecidos do miocárdio de rato em cada grupo; E O número de células TUNEL positivas em cada grupo. F A expressão de mRNA de α-SMA em tecidos do miocárdio de rato em cada grupo foi detectada por RT-qPCR; G A expressão de mRNA de FSP-1 e E-caderina em tecidos do miocárdio de rato em cada grupo foi detectada por RT-qPCR. * P <0,05; ** P <0,01; **** P <0,0001
Discussão
A miocardite é uma doença cardíaca inflamatória que resulta em DCM e insuficiência cardíaca e é mais frequentemente induzida por infecções virais, como CVB3 [2]. Um estudo revelou que o miR-133 alivia a apoptose dos cardiomiócitos e a remodelação elétrica em camundongos com VMC [23]. Além disso, miRNAs exossômicos alterados também estão relacionados à patogênese da miocardite induzida por CVB3 [14]. Exossomos derivados de células progenitoras cardíacas facilitam a apoptose induzida por CVB3 por meio da restrição da proliferação de CVB3 em VMC [24]. Este estudo explorou o mecanismo regulatório do miR-133 exossômico derivado de BMSC na fibrose miocárdica e EMT em ratos VMC (arquivo adicional 1:Fig. 1).
O estudo descobriu que a expressão do miR-133a foi diminuída no VMC. Como demonstrado antes, a expressão do miR-133a está diminuída no MI [7]. Um estudo também sugeriu que a expressão relativa de miR-133 em corações de camundongos do VMC é obviamente diminuída em contraste com os controles [23]. Existem algumas conexões de miRNAs com exossomos. A expressão diferencial de exossomos e de miRNAs exossômicos na doença tem sido considerada como biomarcadores da doença com desempenho de diagnóstico clínico não invasivo juntamente com seus potenciais terapêuticos [25]. Lin et al . descobriram que miR-133 é especialmente classificado em exossomos derivados de células progenitoras endoteliais humanas causadas por hipóxia / reoxigenação (H / R) para aumentar a angiogênese de fibroblastos e EMT [26]. Outro estudo revelou que as MSCs apresentam comunicação com as células do parênquima cerebral e podem modular o crescimento de neuritos pela transferência de miR-133b para células neurais via exossomos [27].
O principal achado deste trabalho manifestou que miR-133a exossômico regulado positivamente promoveu a viabilidade celular, inibiu a inflamação, apoptose, EMT e fibrose em ratos com VMC. Eles se encaixam bem com uma pesquisa anterior de que o silêncio do miR-133a reverte os polissacarídeos do astrágalo inibição da proliferação de células do osteossarcoma MG63 induzida por tratamento, juntamente com a promoção da apoptose celular [28]. Outro estudo revelou que o miR-133a superexpresso suprime a angiogênese, apoptose, fibrose e inflamação, enquanto acelera a remodelação cardíaca terapêutica em doenças isquêmicas do miocárdio [29]. Semelhante ao nosso estudo, Li et al . afirmaram que o miR-133 inibe a apoptose de cardiomiócitos por meio da regulação da expressão de genes relacionados à apoptose no coração de camundongos VMC [23]. Foi relatado que o miR-133a superexpressado deprime a apoptose induzida por hipóxia e fortalece a sobrevivência dos cardiomiócitos [30]. Enquanto isso, os miR-30a e miR-181d exossômicos séricos regulados positivamente podem ter potencial para serem aplicados como biomarcadores para o diagnóstico de VMC [14].
Outro achado em nosso estudo foi que miR-133a exossômico regulado positivamente diminuiu a CVF, reduziu a expressão de colágeno I e colágeno III em ratos com VMC. Um artigo elucidou que o fator de crescimento de fibroblastos 18 liberado de uma membrana de colágeno causa atividade osteoblástica participando de miR-133a regulado negativamente [31]. A expressão excessiva in vitro de miR-133a deprime a hipertrofia dos cardiomiócitos e reduz a expressão do colágeno [32], conforme evidenciado em outro estudo. CVF é igual à razão da área de colágeno para a soma da área do miocárdio e área de colágeno, e o valor médio mostra a CVF da seção [33]. Este achado também é relatado por Wang et al . que o modelo de camundongos VMC é construído com sucesso pela infecção por CVB3, manifestando expressão aparente de CVF mais alta em contraste com o grupo de controle [34]. Além disso, a descoberta é consistente com a de Ferreira et al . que demonstra que o miR-133a pode assumir um papel importante na modulação da expressão gênica na patogênese da cardiomiopatia crônica da doença de Chagas, com potencial link como ferramentas diagnósticas e prognósticas [8]. Além disso, a evidência mostrou que derrubar MAML1 pode reduzir a hipertrofia de cardiomiócitos pré-tratados [35]. Em nosso estudo, descobrimos que MAML1 era o gene alvo de miR-133a e a inibição de MAML1 reverteu os efeitos de exossomos silenciados por miR-133a em ratos com VMC. Na lesão de isquemia-reperfusão miocárdica, a regulação negativa de MAML1 mediada por miR-193b poderia reduzir em parte o infarto e as enzimas miocárdicas, bem como atenuar a apoptose dos cardiomiócitos [18]. Além disso, há um relatório sugerindo que a deficiência de MAML1 pode aliviar a fibrogênese hepática [19].
Conclusão
Em conclusão, o presente estudo oferece evidências de que miR-133a é regulado negativamente em ratos com VMC, e miR-133a exossômico elevado melhora a função cardíaca e restringe a fibrose miocárdica e EMT em ratos com VMC, bem como aumenta a viabilidade e reprime a apoptose de cardiomiócitos em VMC através do direcionamento de MAML1. Nosso estudo também sugere que a inibição de MAML1 tem um efeito protetor em ratos com miocardite e reverte o efeito de exossomos inibidos por miR-133a em ratos com VMC. A identificação do miR-133a exossômico na fibrose miocárdica e EMT da miocardite pode potencialmente ampliar nossa compreensão dos mecanismos subjacentes à miocardite e também ter valor clínico como um novo alvo molecular. Mais pesquisas devem ser realizadas para fazer incursões no tratamento desta doença.
Abreviações
- miR-133:
-
MicroRNA-133
- BMSC-Exo:
-
Exossomo derivado de células-tronco mesenquimais da medula óssea
- EMT:
-
Transição epitelial-mesenquimal
- VMC:
-
Miocardite viral
- CVF:
-
Fração de volume de colágeno
- DCM:
-
Cardiomiopatia dilatada
- CVB3:
-
Vírus Coxsackie B3
- miRNAs:
-
MicroRNAs
- MI:
-
Infarto do miocárdio
- SPF:
-
Livre de patógenos específicos
- SD:
-
Sprague – Dawley
- DMEM:
-
Meio Eagle Modificado por Dulbecco
- PBS:
-
Salina tamponada com fosfato
- FBS:
-
Soro fetal bovino
- NC:
-
Controle negativo
- MOI:
-
Multiplicidade de infecção
- LVPW:
-
Espessura da parede posterior do ventrículo esquerdo
- LVIDs:
-
Diâmetro sistólico final do ventrículo esquerdo
- LVEF:
-
Fração de ejeção do ventrículo esquerdo
- HE:
-
Hematoxilina-eosina
- TÚNEL:
-
Desoxinucleotidil transferase terminal mediada por desoxiuridina trifosfato-biotina nick end-labelin
- ELISA:
-
Ensaio de imunoabsorção enzimática
- TMB:
-
Tetrametilbenzidina
- TBS:
-
Solução salina tamponada com Tris
- ABC:
-
Complexo Avidina-Biotina-Peroxidase
- OD:
-
Densidade ótica
- RT-qPCR:
-
Reação em cadeia da polimerase quantitativa com transcrição reversa
- GAPDH:
-
Gliceraldeído-3-fosfato desidrogenase
- CTGF:
-
Fator de crescimento do tecido conjuntivo
- CCK:
-
Kit de contagem de células
- PI:
-
Iodeto de propídio
- ANOVA:
-
Análise de variação
Detector de ampla faixa de efeito de carregamento induzido por plasma para processos CMOS BEOL avançados
Efeitos da energia acumulada na formação de nanopartículas na reidratação por laser pulsado de filmes finos AgCu
Nanomateriais
- Nanopartículas de fósforo preto promovem a diferenciação osteogênica de EMSCs por meio da expressão de TG2 regulada positivamente
- Estrutura e propriedades eletrônicas da nanoargila caulinita dopada com metal de transição
- Fibras de carbono ativado “Thickly Overgrown” por Ag Nanohair por meio de automontagem e recozimento térmico rápido
- Estudo da Força de Adesão e Transição de Vidro de Filmes de Poliestireno Fino por Microscopia de Força Atômica
- Estudos de TEM e STEM nas morfologias transversais de filmes anti-reflexo de SiO2 de banda larga dupla / tripla
- Uma análise comparativa in vivo de nanopartículas de óxido de cobre e zinco biossintetizadas por vias de administração intraperitoneal e intravenosa em ratos
- Desenvolvimento e caracterização de compostos de cerâmica de vidro contendo Sr com base em hidroxiapatita biogênica
- Materiais da família de grafeno na regeneração do tecido ósseo:perspectivas e desafios
- Nanoscopia baseada em microscopia de força atômica de células-tronco humanas derivadas de adiposo diferenciando condrogenicamente:nanoestrutura e expressão β1 de integrina
- Células-tronco mesenquimais marcadas com nanopartículas de azul da prússia:avaliação da viabilidade celular, proliferação, migração, diferenciação, citoesqueleto e expressão de proteínas …