


Nanopartículas carregadas com hGC33 e carregadas com sorafenib têm um efeito anti-heptatoma sinérgico ao inibir a via de sinalização Wnt
Resumo
A entrega de inibidores específicos do tumor é um desafio no tratamento do câncer. Nanopartículas modificadas com anticorpos podem entregar seus medicamentos carregados às células tumorais que superexpressam antígenos associados a tumores específicos. Aqui, nós construímos nanopartículas de polímero de polietilenoglicol-b-PLGA carregadas com sorafenibe modificadas com anticorpo hGC33 para glypican-3 (GPC3 +), uma proteína de membrana superexpressada em carcinoma hepatocelular. Descobrimos que NPs modificados por hGC33 (hGC33-SFB-NP) direcionados a GPC3 + As células do carcinoma hepatocelular (HCC) ligando-se especificamente a GPC3 na superfície das células HCC, inibiram a transdução de sinal induzida por Wnt e inibiram as células HCC em G0 / 1 pela regulação negativa da expressão de ciclina D1, atenuando assim a migração de células HCC pela inibição epitelial - transição mesenquimal. O hGC33-SFB-NP inibiu a migração, progressão do ciclo e proliferação de células HCC ao inibir a via Ras / Raf / MAPK e a via Wnt em tandem com moléculas GPC3, respectivamente. hGC33-SFB-NP inibiu o crescimento do câncer de fígado in vivo e melhorou a taxa de sobrevivência de camundongos com tumor. Concluímos que hGC33 aumenta o direcionamento de SFB-NP para células HCC. hGC33-SFB-NP inibe sinergicamente a progressão de HCC, bloqueando a via Wnt e a via Ras / Raf / MAPK.
Introdução
Glypican3 (GPC3) é um proteoglicano de heparan sulfato expresso na superfície celular por meio de um mecanismo que envolve âncoras de glicerofosfatidilinositol [1]. Embora GPC3 seja expresso em uma variedade de tecidos durante o desenvolvimento, sua expressão é pelo menos parcialmente inibida na maioria dos tecidos adultos pela metilação do DNA na região do promotor [2]. No entanto, a proteína GPC3 é superexpressada em cerca de 70% dos pacientes com carcinoma hepatocelular (HCC) [3, 4] e estimula a transdução do sinal Wnt clássico [5] por meio da interação com o ligante Wnt, que promove a ligação Wnt / frizzled para promover o crescimento do HCC [6] . A ativação da via clássica de sinalização Wnt é um dos eventos mais frequentes associados à transformação maligna do CHC [7, 8]. Com base na capacidade do GPC3 de aumentar a sinalização Wnt, formulamos a hipótese de que a superexpressão de GPC3 promove o crescimento do HCC ao estimular a via Wnt clássica.
O anticorpo monoclonal terapêutico que reconhece um epítopo na porção C-terminal de GPC3 (524-563) (hGC33), que reconhece o epítopo C-terminal de GPC3 (524-563), inibe o crescimento tumoral em xenoenxertos subcutâneos de HepG2 e Huh- 7 em camundongos [9, 10]. O hGC33 foi humanizado pelo transplante complementar da região de decisão e seu efeito anticâncer é tão eficaz quanto o hGC33 para o xenotransplante de HepG2, e o hGC33 foi usado em ensaios clínicos [11]. Estes resultados sugerem que hGC33 tem importante atividade antitumoral, e o tratamento anti-GPC3 inibirá diretamente a proliferação e / ou sobrevivência de células HCC, bloqueando Wnt e / ou outras vias de sinalização.
No entanto, devido à complexa patogênese do CHC, a eficácia do bloqueio de GPC3 sozinho é limitada [12]. Portanto, explorar o mecanismo detalhado da patogênese do CHC e identificar biomarcadores promissores para o diagnóstico e prognóstico do CHC pode ajudar a fornecer alvos terapêuticos eficazes e melhorar o prognóstico dos pacientes. Os anticorpos anti-GPC3 foram propostos para aumentar a sensibilidade do HCC aos agentes quimioterápicos [13]. A atividade antitumoral de hGC33 em combinação com drogas quimioterápicas padrão foi avaliada recentemente [14]. O sorafenibe é um novo medicamento de HCC direcionado por via oral que pode inibir diretamente a proliferação de células tumorais, bloqueando a via de sinalização celular mediada por RAF / MEK / ERK para conter o crescimento tumoral [15, 16]. No entanto, a distribuição não direcionada de sorafenibe in vivo e sinais Wnt anormalmente ativados no HCC limitam a eficácia da droga e aumentam seus efeitos colaterais [14,15,16]. Wnt se liga a receptores na família Frizzled para ativar a transdução de sinal intracelular que regula a proliferação celular, apoptose e migração celular, e causa resistência aos medicamentos em muitos tumores, como o CHC [17,18,19].
Em modelos de xenotransplante de HepG2, a combinação de hGC33 e sorafenibe é mais eficaz na inibição do crescimento tumoral do que o sorafenibe sozinho [15], e a distribuição de drogas com nanocarreadores de polímero tem recebido muita atenção no tratamento do câncer. Os nanocarreadores carregados de drogas anticâncer podem prevenir a distribuição inespecífica e a degradação não específica in vivo, melhorar a biodisponibilidade da droga e o direcionamento antitumoral e simplificar a avaliação da farmacocinética e do tratamento [15, 16]. Em uma variedade de nanopartículas à base de polímero, as formulações à base de poli (ácido lático-glicólico) (PLGA) são consideradas transportadoras de drogas ideais e seguras [17, 18]. A este respeito, o poli (etilenoglicol) - b -poly (d, l-lactídeo-co-glicolídeo) (PEG- b -PLGA) são baseados em polietilenoglicol e copolímeros de PLGA, que são seguros e não tóxicos após hidrólise, e foram aprovados pela US Food and Drug Administration [19,20,21,22]. Portanto, a injeção intravenosa com nanocarreador de PEG- b O copolímero -PLGA é uma estratégia promissora para alcançar a entrega direcionada e aumentar a eficácia. Além disso, a aplicação do anticorpo específico hGC33 contra moléculas de GPC3 na membrana da célula HCC pode não apenas melhorar a distribuição de nanofármacos direcionados in vitro e in vivo [18, 19], mas também bloquear Wnt e / ou outras vias de sinal conectadas com GPC3, inibir a proliferação e / ou sobrevivência de células cancerosas e pode atingir atividade antitumoral sinérgica.
Neste estudo, exploramos se o copolímero modificado com anticorpo hGC33 PEG- b Nanopartículas de -PLGA podem facilitar a entrega de sorafenibe (hGC33-SFB-NP) ao HCC in vivo e in vitro e melhorar a eficiência do tratamento de HCC por meio do direcionamento ativo de HCC para alterar a farmacocinética do medicamento. De acordo com o tamanho da partícula, potencial zeta, morfologia da partícula, eficiência de aprisionamento da droga, capacidade de carga da droga e liberação da droga in vitro, o NP alvo foi amplamente caracterizado. A capacidade de direcionamento in vitro é caracterizada pela absorção celular de células de hepatoma HepG2. A biodistribuição e o efeito terapêutico sinérgico de hGC33-SFB-NP em HCC foram avaliados comparando sorafenibe e SFB-NP. Nossos resultados demonstraram que hGC33-SFB-NP pode ter como alvo GPC3 + HCC. Ele pode inibir a progressão do ciclo celular, a proliferação celular e a invasão tumoral ao inibir as vias de sinal Wnt e Ras / Raf / MAPK e inibir sinergicamente a progressão do câncer de fígado.
Materiais e métodos
Materiais
O anticorpo hGC33, brometo de 3- (4,5-dimetiltiazol-2-il) -2,5-difeniltetrazólio (MTT), 4 ′, 6-diamidino-2-fenilindol, dicloridrato (DAPI), 5,5 ′, 6 , Iodeto de 6′-tetracloro-1,1 ′, 3,3′-tetraetil-benzimidazolil carbocianina (JC-1) e dimetilsulfóxido (DMSO) foram obtidos de Sigma-Aldrich, Inc. (St. Louis, MO). Cloridrato de 1- (3-dimetilaminopropil) -3-etil-carbodiimida e N -hidroxissuccinimida foram obtidas na Qiyun Biotech (Guangzhou, China). O kit de quantificação de proteína de ácido bicinconínico (BCA), cumarina-6 e o kit de detecção de apoptose de Anexina V-FITC / PI foram adquiridos na Beyotime Biotechnology (Shanghai, China). PEG- b -PLGA-copolímero dibloco de maleimida (mal-PEG- b -PLGA; 25.000-30.000 Da, PLGA, LA:GA, w / w; PEG, 13–15%) foi adquirido da Polyscitech (West Lafayette, IN, EUA). Kit de anticorpo PI3K (9655 #), kit de anticorpo p-Akt (9916 #), kit de anticorpo mTOR (9964 #), kit de anticorpo da família Bcl-2 (9942 #), kit de anticorpo de apoptose (9915 #) e anti- os anticorpos de coelho e anti-rato foram adquiridos na Cell Signal Technology (Danvers, MA, EUA); a ciclina B1 e a quinase dependente de ciclina foram adquiridas na Abcam Biological Technology (EUA). Os anticorpos para fosfo-Rb, ciclina D1, checkpoint quinase 1 (CHK1), P53, gene de susceptibilidade ao câncer de mama fosforilado 1 (p-BRCA1), RAD51, citocromo C e metaloproteinase de matriz (MMP2 e MMP9) foram adquiridos da Abcam (EUA) . Todos os outros produtos químicos de grau analítico, reagentes e solventes são obtidos de fornecedores padrão e foram usados sem purificação adicional.
Células e animais
A linha de células HCC HepG2 (obtida da coleção de cultura americana (Manassas, VA, EUA) foi cultivada em meio DMEM (Invitrogen, Carlsbad, CA, EUA) suplementado com 10% de FBS (HyClone, Logan, UT, EUA) e 80 Penicilina U / ml e estreptomicina 80 μg / ml em uma atmosfera umidificada de 5% de CO 2 a 37 ° C. Camundongos pelados BALB / C, pesando 20–22 g (5–6 semanas) foram fornecidos por Nanjing Junke Biotechnology Co., Ltd. (China). Camundongos nus BALB / C foram criados em uma sala SPF. Todos os cuidados e tratamentos dos animais foram realizados de acordo com os requisitos de cuidados com os animais da Universidade de Ciência e Tecnologia de Anhui. Todos os protocolos experimentais foram revisados e aprovados pelo comitê de ética em experimentos com animais da Universidade de Ciência e Tecnologia de Anhui (Nº de aprovação:2019dw013).
Preparação de NPs
Para preparar NPs, mal-PEG- b -PLGA e SFB ou cumarina-6 foram pesados e dissolvidos na fase orgânica (3:2 v / v diclorometano / acetona). A solução foi adicionada a uma solução de álcool polivinílico (PVA) (5% p / v) gota a gota com agitação contínua. A mistura foi sonicada intermitentemente com um sonicador de sonda (potência de saída de 550 W, 8 vezes) em gelo para criar uma emulsão de óleo-água. A emulsão foi adicionada a uma solução de PVA (1% p / v) com agitação magnética. SFB e NPs de cumarina-6 foram coletados por centrifugação a 8.000 rpm por 30 min e lavados três vezes em água Milli-Q.
Para gerar hGC33-NP por ligações tioéter formadas pela reação de maleimida com resíduos de sulfidrila livres no anticorpo hGC33, o anticorpo hGC33 foi misturado com NPs funcionalizados com maleimida a uma razão molar de 5:1 (hGC33:mal-PEG- b -PLGA) e incubado a 4 ° C por 16 h com agitação contínua. O hGC33 foi conjugado aos NPs através da reação de grupos sulfidrila no anticorpo hGC33 com os grupos maleimida das cadeias de PEG. Os anticorpos não conjugados foram removidos por passagem através de colunas Sepharose CL-4B. A conjugação eficiente de proteínas foi confirmada com um kit BCA (Thermo Fisher Scientific, Waltham, MA, EUA).
Caracterização de Nanopartículas (NPs)
Morfologia, tamanho de partícula, eficiência de encapsulamento (EE) e estabilidade de NPs
A morfologia dos NPs foi avaliada com microscopia eletrônica de transmissão (TEM, H-600; Hitachi, Tóquio, Japão). HGC33-NPs livres de drogas e NPs livres de drogas foram registrados por espectrofotômetro FTIR (Thermo Nicolet, Madison, WI, EUA) usando brometo de potássio. O tamanho médio de partícula e o potencial zeta de NPs foram caracterizados com um Malvern Zetasizer ZEN3600 Nano ZS (Malvern Instruments, Malvern, Reino Unido) a 20 ° C. A eficiência de encapsulação da droga (EE) e a eficiência do conteúdo de carga da droga (LC) foram avaliadas por ultrafiltração. As amostras foram carregadas em um dispositivo de ultrafiltração (100 kMWCO; Sartorius, Göettingen, Alemanha) e centrifugadas a 8.000 rpm por 25 min a 4 ° C para remover a droga livre. O mesmo volume de cada amostra foi dissolvido em acetonitrila para confirmar a quantidade total de fármaco. A concentração foi medida por cromatografia líquida de alta eficiência. O comprimento de onda de absorção foi de 266 nm. A seguinte fórmula foi usada para calcular o EE do fármaco (%) dos NPs:(peso do fármaco aprisionado / peso total do fármaco) × 100%. LC (%) foi calculado como (peso do fármaco encapsulado / peso de NPs) × 100%. Para compreender a estabilidade dos NPs à temperatura ambiente, a mudança no tamanho do NP foi avaliada por espalhamento dinâmico de luz (DLS) em pontos de tempo predeterminados (0,5, 1, 2, 4, 8, 12 h, 16 h, 20 h e 24 h) a 25 ° C.
Liberação de droga in vitro e ensaio de absorção celular
A droga dos NPs foi investigada usando bolsas de diálise com um peso molecular de corte de 10 kDa. Resumidamente, 1 mL de NPs foi carregado em uma bolsa de diálise (MWCO 8.000–10.000 Da; Spectrum Labs Inc., CA, EUA). Os sacos de diálise foram imersos em PBS e agitados com um agitador magnético a 25 ° C. Os perfis de liberação de drogas de NPs foram medidos em 100 mL de solução salina tamponada com fosfato 0,2 M (PBS; pH =7,4) por 7 dias. A concentração do fármaco nas amostras foi medida por cromatografia líquida de alta eficiência. Em estudos subsequentes, hGC33-cumarina 6-NP com o mesmo tamanho de partícula que hGC33-SFB-NP foi usado para avaliar o direcionamento de hGC33-SFB-NP. HepG2 (GPC3 + ) e Li-7 (GPC3 - ) foram incubados com hGC33-cumarina 6-NP por 0,5, 2 ou 4 h a 37 ° C em 5% de CO 2, respectivamente. As células co-cultivadas foram lavadas e fixadas com formaldeído 4% por 10 min; os núcleos das células foram corados com 5 μg / mL Hoechst 33.342 por 15 min para localizar NPs intracelulares. Microscopia confocal para analisar imagens de nanopartículas intracelulares foi analisada com microscopia confocal (Olympus FV1000; Olympus Corporation, Tóquio, Japão).
Efeito celular in vitro
A citotoxicidade de hGC33 (Ab) livre, SFB livre, hGC33-null-NP ou hGC33-SFB-NP foi determinada usando um ensaio de MTT. HepG2 (GPC3 + ) células e Li-7 (GPC3 - ) células em fase logarítmica foram semeadas em uma placa de 96 poços a uma densidade de 4000 células por poço, seguido por incubação por 48 h a 37 ° C em 5% de CO 2 . As células foram tratadas com hGC33, SFB livre, hGC33-null-NP ou hGC33-SFB-NP por 48 h a 37 ° C em 5% de CO 2 . Após o co-cultivo por um tempo definido, a atividade anti-proliferação celular foi determinada pelo ensaio MTT conforme descrito [20]. A absorvância de cada poço foi medida a 490 nm e calculada a metade do valor da concentração inibitória máxima (IC50) com SPSS 17.0.
Medição da capacidade de invasão celular
As células na fase de crescimento logarítmico foram semeadas em placas de 6 poços a uma densidade de 5 × 10 4 células / poço, riscado com uma cabeça de sucção e substituído por meio de cultura sem soro. A cicatrização da ferida foi registrada em 0 h, 24 h e 48 h no grupo controle e no grupo experimental. Ao mesmo tempo, o mesmo número de células foi inoculado em câmaras Transwell e tratado com hGC33 livre, SFB livre, hGC33-null-NP ou hGC33-SFB-NP. Quinhentos microlitros de meio FBS a 10% foram adicionados à câmara inferior. Após incubação por 24 h, a câmara Transwell foi retirada e as células foram fixadas com paraformaldeído a 4% e coradas com violeta de cristal 0,1%. O tamanho da cicatrização da ferida e o número de células de migração foram calculados para avaliar a capacidade de migração.
Determinação do ciclo celular
Após incubação durante a noite em placas de 6 poços, as células foram tratadas com hGC33 livre (Ab), SFB livre, hGC33-null-NP ou hGC33-SFB-NP por 24 h, depois coletadas e fixadas com etanol. Após a coloração com iodeto de propídio, foi realizada citometria de fluxo e o ciclo celular foi analisado com modifit 3.0 (Verity Software House, Topsham, ME).
Western Blotting
Para avaliar o status de ativação da via de sinal e a expressão de moléculas alvo, as células foram incubadas durante a noite em placas de 6 poços, e hGC33 livre, SFB livre, hGC33-null-NP ou hGC33-SFB-NP livre foram aplicados por 24 h . As células de cada grupo de tratamento foram coletadas e a proteína foi extraída e medida. A concentração de proteína foi medida e calibrada com o kit de proteína BCA (Biosharp, Hefei, China). A proteína das amostras foi separada por doze eletroforese em gel de alquilsulfato poliacrilamida, transferida para uma membrana de PVDF e selada com leite desnatado. O primeiro anticorpo (diluído 1:1000) foi incubado durante a noite a 4 ° C, e os segundos anticorpos (1:2000) foram incubados por 1 h em temperatura ambiente. As bandas foram visualizadas com substratos ECL (Thermo Fisher Scientific Waltham, MA, EUA), e as imagens foram visualizadas em sistema de análise de imagens em gel, sendo a β-actina usada como controle.
Atividade antitumoral in vivo
Foi determinada a inibição de hGC33 livre, SFB livre, hGC33-null-NP, SFB-NP e hGC33-SFB-NP no crescimento de HCC in vivo. De acordo com os regulamentos e diretrizes sobre saúde animal do comitê de ética da Universidade de Tecnologia de Anhui, todos os experimentos foram conduzidos em camundongos BALB / c em gaiolas em uma sala de controle de temperatura (23 ± 2 ° C) com 12 h / 12 h de luz / ciclo escuro. Uma suspensão de 50 μl contendo 5 × 10 6 células HepG2 vivas foram injetadas por via subcutânea no abdômen direito de camundongos BALB / c fêmeas de 5 semanas de idade (20-22 g). Quando o volume do tumor atingiu cerca de 50 mm 3 , os ratos foram divididos aleatoriamente em 6 grupos (10 ratos em cada grupo). Controle salino normal NS (200 mg / kg NP nulo em 200 μL de PBS), hGC33-null-NP (hGC33-null-NP em 200 μL de PBS, equivalente a hgc33 =100 μg / kg / tempo), hGC33 livre (hGC33 em 200 μL de PBS, 100 μg / kg / tempo), SFB livre (dose de SFB:8 mg / kg / tempo), SFB-NP (dose de SFB:8 mg / kg / tempo) e hGC33-SFB-NP (equivalente a SFB =8 mg / kg / tempo, hGC33 =100 μg / kg / tempo) foram injetados na veia da cauda a cada 2 dias por 10 vezes. O peso e o tamanho do tumor dos camundongos foram medidos a cada quatro dias. A fórmula de cálculo do volume do tumor foi volume =0,5 × L × W 2 , onde L e W representam o comprimento e a largura do tumor, respectivamente. Quatro semanas após a administração, os animais foram anestesiados com éter dietílico e mediram-se o tamanho e o peso do tumor. Além disso, tumor, coração, fígado, rim, pulmão e baço foram removidos, fixados com solução de paraformaldeído a 4%, incluídos em parafina, seccionados e corados com hematoxilina e eosina para avaliação das alterações histológicas por microscopia digital.
Análise estatística
Os dados são apresentados como média ± desvio padrão (DP) e foram avaliados por análise de variância com SPSS 18.0. Comparações estatísticas pareadas foram realizadas usando um teste t de Student bicaudal. As diferenças foram consideradas estatisticamente significativas para P <0,05.
Resultados
Caracterização de NPs e liberação de drogas In Vitro
Como o diâmetro e as propriedades de superfície do NP afetam a absorção celular, a liberação do fármaco e a distribuição do NP in vivo, caracterizamos o NP preparado com os parâmetros correspondentes. A morfologia, tamanho de partícula e distribuição de tamanho de partícula dos polímeros SFB-NP, hGC33-null-NP e hGC33-SFB-NP estão resumidos na Fig. 1. A morfologia de hGC33-SFB-NP observada por microscópio eletrônico de transmissão (Fig. . 1a) mostra núcleos rígidos com bordas difusas, o que indica que hGC33 está presente na superfície de NPs. Os tamanhos de partícula de SFB NP, hGC33-null-NP e hGC33-SFB-NP variaram de 100 a 150 nm e tinham distribuição de tamanho de partícula unimodal típica. O diâmetro médio de hGC33-SFB-NP (120,2 ± 10,2 nm) foi ligeiramente maior do que o de hGC33-null-NP, SFB-NP e NP nulo (Fig. 1a, Tabela 1). Filmes esféricos finos não claros com uma única superfície estavam presentes na superfície de hGC33-SFB-NP e hGC33-null-NP, o que indicava que o anticorpo hGC33 estava presente na superfície de NPs. O tamanho aumentado para hGC33-SFB-NP e hGC33-null-NP confirmou a existência do filme hGC33. A síntese de hGC33-SFB-NP foi confirmada com 1 H-NMR (Fig. 1b) antes da preparação de NP. Os picos a 5,2 ppm e 1,58 ppm foram atribuídos a prótons -CH3 do ácido lático; o pico a 4,8 ppm foi atribuído a -OCH2- do ácido glicólico; e o pico em 3,6-3,8 ppm foi atribuído aos prótons -CH2CH2O- de unidades de repetição de PEG. Química de superfície de PEG- b -PLGA e Ab-PEG- b -PLGA NPs também foi estudado por espectroscopia FTIR (Fig. 2c). No espectro de PEG- b -PLGA polímero, uma banda forte em cerca de 1750 cm −1 originado do trecho dos grupos carbonila (C =O) da cadeia do PLGA. Uma banda em 2880 cm −1 foi devido ao alongamento de um grupo a –CH na cadeia de PEG. Ao mesmo tempo, um pico apareceu em 2520 cm −1 que atribuímos ao pico de alongamento -SH do anticorpo hGC33. O 1 Os resultados de H NMR e FTIR indicaram que o anticorpo foi enxertado na estrutura do PEG- b Polímeros -PLGA.
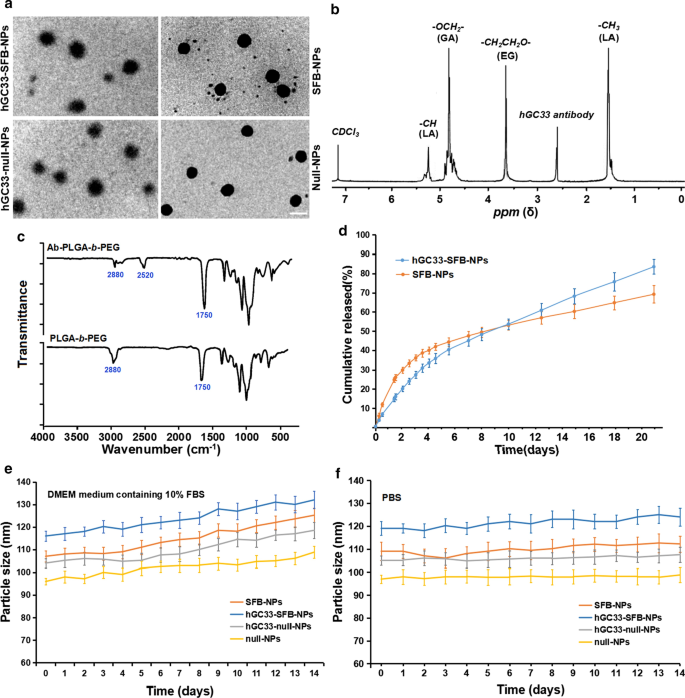
Caracterização de NPs. a Caracterização TEM de NPs, a barra de escala indica 100 nM; b o 1 Espectro de H NMR de hGC33-PLGA- b sintético -PEG em CDCl3; c Espectros de FTIR de hGC33-PEG- b -PLGA e PEG- b - PLGA; d perfis de liberação cumulativa de SFB-NP e hGC33-SFB-NP em PBS (pH =7,4) a 37 ° C; e mudanças de tamanho de NPs incubados em meio DMEM contendo 10% de FBS ao longo de 14 d; f mudanças de tamanho de NPs incubados em PBS ao longo de 14 d. SFB, sorafenib; TEM, microscopia eletrônica de transmissão; NPs, nanopartículas; 1 H NMR, 1 Espectroscopia de ressonância magnética nuclear H; FTIR, espectroscopia de infravermelho com transformada de Fourier
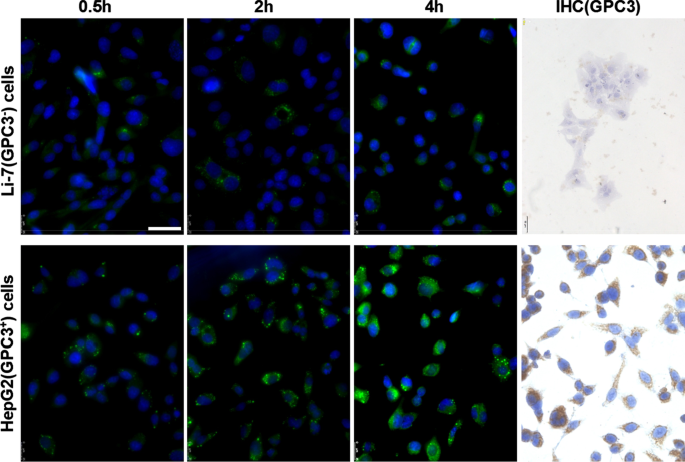
Expressão de GPC3 e absorção de hGC33-cumarin6-NP em células Li-7 e HepG2. As células, inoculadas na placa de cultura, foram lavadas com PBS e incubadas com 100 μg / ml de hGC33-cumarin6-NP em DMEM por 2, 4 e 8 h. Os núcleos foram corados com DAPI e as células foram fixadas e detectadas por microscopia confocal de varredura a laser. GPC3 não foi detectado em células Li-7, mas foi altamente expresso em células HepG2 conforme detectado por imunocitoquímica. A barra de escala indica 50 μM
Curiosamente, as nanopartículas de hGC33-SFB-NP e SFB-NP em meio DMEM contendo 10% de FBS (pH =7,4) tiveram liberação rápida do fármaco até cerca de 4 dias, depois liberação do fármaco relativamente lenta e estável; a liberação de SFB cumulativa de hGC33-SFB-NP e SFB-NP ao longo de 20 dias foi de cerca de 77% e 65%, respectivamente (Fig. 1d). A diferença pode ser devido à molécula hidrofílica na superfície do PEG- b Matriz -PLGA, que pode acelerar a degradação das nanopartículas, aumentando a hidratação e, assim, promovendo a hidrólise. Para determinar a estabilidade dos NPs preparados, o hGC33-SFB-NP, hGC33-NP nulo, NP-nulo e SFB-NP foram colocados em meio DMEM contendo 10% de FBS (pH =7,4) e em PBS (pH =7,4 ) Os tamanhos dos vários NPs permaneceram estáveis por mais de 2 semanas. SFB foi liberado de hGC33-SFB-NP de uma maneira sustentada e estável por 14 dias, mas houve uma mudança modesta no tamanho das partículas no meio DMEM em comparação com aquele em 10% de FBS (Fig. 1e, f). A estabilidade de hGC33-SFB-NP seria apropriada para SFB com um papel biológico sustentado.
Grande potencial zeta pode causar forte interação repulsiva eletrostática entre NP e manter a estabilidade do sistema de dispersão NP [23, 24]. Como mostrado na Tabela 1, os potenciais zeta de hGC33-SFB-NP, hGC33-null-NP e SFB-NP são - 18,2 ± 2,2 mv, - 18,5 ± 1,8 mv, e - 15,9 ± 2,1 mv, respectivamente, que podem ser causada pelas cargas negativas geradas pelo grupo aldeído, grupo carboxila e grupo fosfato na glicoproteína do anticorpo hGC33. Os NPs carregados negativamente podem levar à alta estabilidade da suspensão NP. Além disso, como a superfície das células é carregada negativamente em ambientes fisiológicos normais, os NPs preparados repelem as células com baixa carga e são menos tóxicos para os tecidos e células. Além disso, a distribuição de tamanho de null-NP e SFB-NP (índice de polidispersidade [PDI] 0,18 e 0,19, respectivamente) foi ligeiramente, mas não significativamente menor do que a de hGC33-null-NP (PDI =0,21) e hGC33-SFB- NP (PDI =0,23). Assim, o tamanho das nanopartículas tem boa uniformidade. O tamanho de partícula, distribuição de tamanho de partícula e potencial zeta de NP são mostrados na Tabela 1 de acordo com os resultados da eficiência de encapsulação SFB e conteúdo de carregamento. O anticorpo GPC3 não conjugado hGC33 foi removido por ultracentrifugação e a eficiência de ligação do anticorpo hGC33 a NPs foi avaliada. A análise da proteína BCA mostrou que a eficiência de ligação do anticorpo hGC33 a NPs foi de 79,5% ± 2,9%.
hGC33-Coumarin 6-NP Almeja efetivamente GPC3 + Células HepG2 da linha celular HCC
Para descobrir se o anticorpo hGC33 de nanopartículas ainda tem a capacidade de direcionar especificamente GPC3, usamos GPC3 + HepG2 e GPC3 - Células Li-7 como células alvo e hGC33-cumarina 6-NP como nanopartículas traçadoras, e incubou as diferentes células por 2 h, 4 h e 8 h. As células foram lavadas com PBS 3 vezes e reagiram com DAPI para manchar o núcleo. As células foram fixadas e fotografadas com microscópio de fluorescência Leica (DMi8, Alemanha). Verificou-se que a fluorescência verde nas células HepG2 era significativamente maior do que nas células Li-7 no mesmo tempo de incubação (Fig. 2), indicando que a quantidade de hGC33-cumarina 6-NP entrando nas células HepG2 era significativamente maior do que em células Li-7. Os resultados documentaram que o anticorpo hGC33 em hGC33-Coumarin6-NP ainda tinha a capacidade de direcionar GPC3 e mediar a internalização de nanopartículas. A expressão de GPC3 em células HepG2 e células Li-7 foi examinada com fluorescência indireta e coloração citoquímica. Os resultados mostraram que as células HepG2 superexpressaram GPC3, enquanto as células Li-7 não expressaram GPC3 (Fig. 2).
hGC33-Null-NP inibe a proliferação de células HepG2
Para determinar se NP modificado com hGC33 (hGC33-null-NP) poderia inibir o crescimento de HCC, medimos a inibição do crescimento celular da linha de células de HCC positiva para GPC3 HepG2 e da linha de células negativa de GPC3 Li-7. Descobrimos que hGC33-null-NP e hGC33 inibiram o crescimento da linha de células HCC GPC3-positivas HepG2 após 24 h de tratamento, mas hGC33 teve efeito inibidor mais significativo em células HepG2 do que hGC33-null-NP (Fig. 3a); em contraste, hGC33-null-NP e hGC33 não afetaram o crescimento da linha celular Li-7 negativa para GPC3 (Fig. 3b). O curso de tempo representativo de hGC33-null-NP (equivalente a 0,1 μm hgc33) e hGC33 (0,1 μm) na proliferação de células HepG2 é ilustrado na Fig. 3c. Como hGC33 é um peptídeo C-terminal que reconhece GPC3, hGC33 é superior a hGC33-null-NP na inibição de HepG2. Os resultados sugerem que a molécula de hGC33 em hGC33-null-NP pode ter efeito de bloqueio de espaço, o que pode afetar a capacidade de ligação de hGC33 a epítopos.
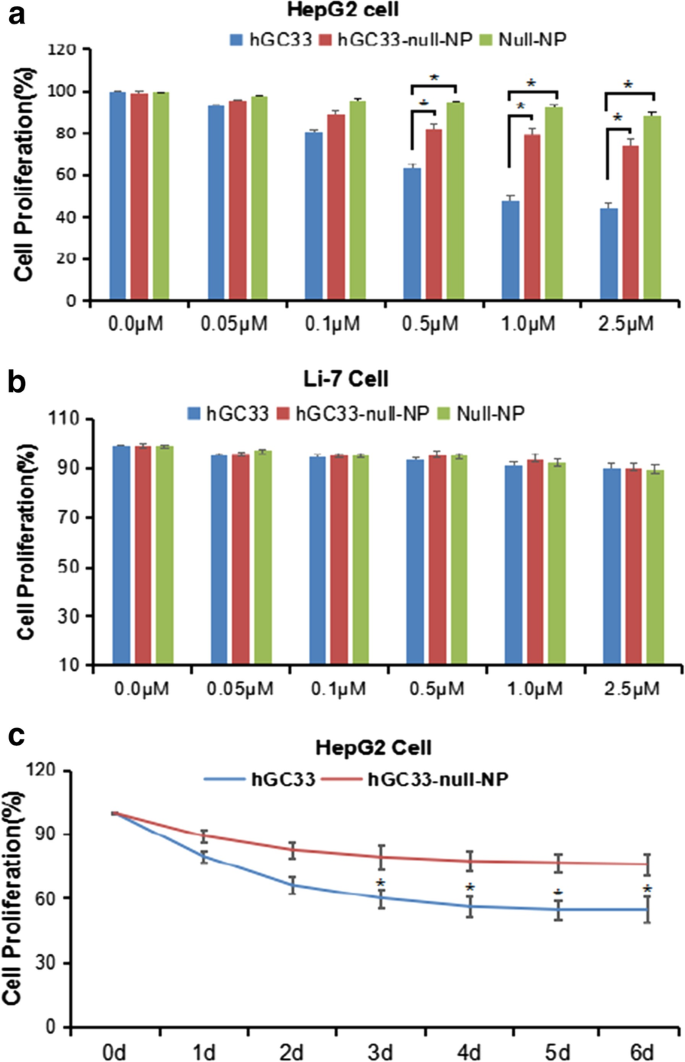
A proliferação de células HepG2 e Li-7 foi inibida por hGC33, nulo-NP e hGC33-nulo-NP. a O teste de proliferação celular foi realizado em células HepG2 positivas para GPC3 tratadas com hGC33, NP-nulo ou NP-nulo de hGC33; b os testes de proliferação celular foram conduzidos em células Li-7 negativas para GPC3 tratadas com hGC33, NP nulo e NP nulo hGC33. As células foram incubadas com 0-2,5 μM hGC33, NP nulo ou NP nulo hGC33 por 1 dia. A proliferação das células foi medida pelo método MTT e padronizada como células não tratadas. Todos os valores representam a média ± DP. Em comparação com o grupo de controle (0 μM) sem tratamento com anticorpos, * P <0,01; c os resultados representativos da resposta de HepG2 a hGC33, null-NP e hGC33-null-NP no tratamento com hGC33 (1,0 μM)
hGC33-Null-NP inibe o ciclo celular das células HepG2
Para entender o mecanismo da atividade molecular do anticorpo hGC33 modificado em nanopartículas de NP nulo de hGC33, estudamos a progressão do ciclo celular de HepG2 tratado com hGC33-NP nulo. Na linha celular HepG2, hGC33-null-NP e hGC33 aumentaram significativamente a proporção de células em G1 (Fig. 4a, b), indicando que hGC33-null-NP e hGC33 poderiam induzir a interrupção do ciclo celular na fase G1. Além disso, hGC33-null-NP e hGC33 podem diminuir significativamente a expressão de ciclinD1 em células HepG2 (Fig. 4c, d).
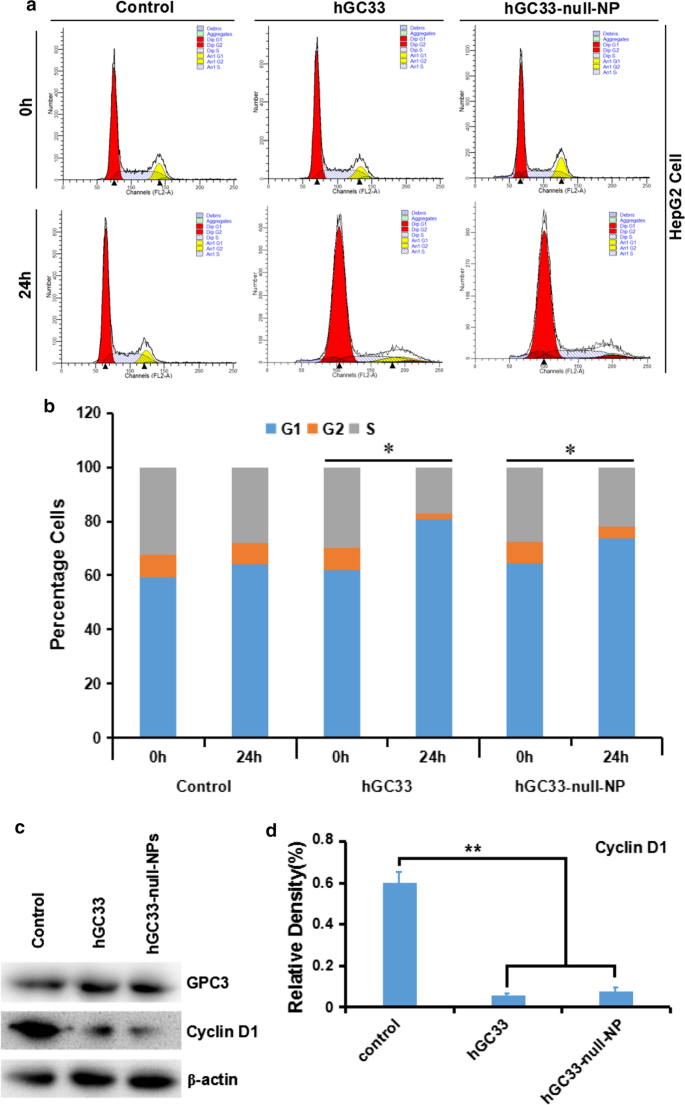
Parada do ciclo celular induzida por hGC33-null-NP e hGC33 na fase G1 e inibição da expressão de ciclinD1 em células HepG2. a Diagrama do ciclo celular representativo de vários grupos tratados com hGC33-null-NP e hGC33. b Análise do ciclo celular de vários grupos tratados com hGC33-null-NP e hGC33. As células HCC foram incubadas com 0,5 μm de hGC33 ou hGC33-null-NP (com a concentração molar de hGC33 como referência). * P <0,05, a fase G1 das células hGC33-null-NP ou hGC33 foi comparada com a das células 0 h. c , d Após o tratamento com hGC33-null-NP ou hGC33, a ciclinD1 foi significativamente regulada para baixo em células HepG2 em comparação com o grupo de controle
Ativação Wnt em células HepG2, Huh-7 e Li-7
Para entender a ativação do sinal Wnt / β-Catenina clássico em células HCC, primeiro detectamos a expressão do ligante Wnt e da proteína crimp do receptor (frizzled, FZD ou Frz) em várias linhas de células HCC:HepG2 (GPC3 ++ ), Huh-7 (GPC + ) e Li-7 (GPC3 - ) Os resultados mostraram que os receptores Wnt3a e FZD de transduzir a via da β-Catenina foram expressos em todas as linhas celulares, especialmente em células HepG2 e Huh-7 (Fig. 5).
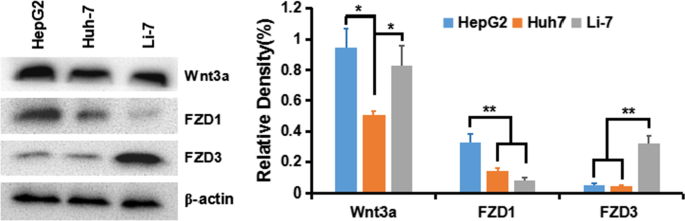
Expressão de Wnt3a, FZD1 e FZD3 em linhas celulares HCC
hGC33-Null-NP inibe a proliferação celular dependente da transdução do sinal Wnt induzida por Wnt3a
Alguns estudos têm demonstrado que a parte extracelular de GPC3 pode ser um co-receptor de Wnt, que promove a ativação e transdução do sinal Wnt / β-Catenina. Portanto, quando HepG2 (GPC3 + ), Huh-7 (GPC3 + ) e Li-7 (GPC3 - ) as células foram co-incubadas com hGC33 e hGC33-null-NP, a ativação do sinal Wnt / β-catenina foi bloqueada por hGC33 e hGC33-null-NP e proliferação de HepG2 e Huh-7, mas não Li-7, in Wnt3a-conditioned medium was inhibited. That proliferation of Li-7 cells most likely is due to the absence of GPC3 on the surface of Li-7 cells (Fig. 6). Our results indicate hGC33 and hGC33-null-NP nanoparticles specifically bind to GPC3 molecules on cell membrane. hGC33 and hGC33-null-NP partially neutralize the mitogenic activity of Wnt3a and inhibit the Wnt/β-catenin signaling pathway. However, the inhibition of proliferation by hGC33-null-NP nanoparticles was less than that of hGC33. Perhaps the spatial structure of the nanoparticles interferes with hGC33-null-NP and limits the function of the hGC33 molecule on the nanoparticles so they cannot completely block the interaction between GPC3 and Wnt3a.

GPC3-activated Wnt signal transduction in HCC. Fifty percent Wnt3a DMEM medium was added with anti-wnt3a antibody, hGC33, or hGC33-null-NP. HepG2 (GPC3 ++ ), Huh-7 (GPC3 + ) and Li-7 (GPC3 − ) cells were co-incubated for 48 h, and cell proliferation was measured by MTT assay. The data were expressed as mean ± SD (*P < 0.01)
hGC33-Null-NP Inhibits Wnt3a-Induced Signal Transduction in HepG2 and Huh-7 Cells
To understand the effect of hGC33-null-NP on Wnt/β-catenin signaling in HCC cells, we extracted the proteins of HepG2 and Huh-7 cells treated with hGC33 or hGC33-null-NP. The results of western blot showed that after hGC33-null-NP treatment, the levels of pYAP and pβ-catenin were increased, but the levels of YAP and β-catenin were decreased. Furthermore, the levels of cyclinD1, CD44, VEGF, and c-MYC in the hGC33-null-NP group were lower than those in the control group, but the level was less than with hGC33 treatment. Similar effects were observed in HepG2 and Huh-7 cells, as shown in Fig. 7.

Inhibition of Wnt3a-induced β-catenin signaling by hGC33-null-NP or hGC33. a Compared with the control group, Wnt/β-catenin signaling pathway in HepG2 and Huh-7 cells treated with hGC33-null-NP or hGC33 was inhibited, and the levels of β-catenin and YAP were decreased, while the increased phosphorylated β-catenin and phosphorylated YAP molecules were unstable, and degraded in cytoplasm. The decreased β-catenin was difficult to maintain in the nucleus and drive the expression of CyclinD1, CD44, VEGF, and c-MYC, which resulted in the decrease of cyclinD1, CD44, VEGF, and c-myc protein levels. b The mechanism pattern of Wnt/β-catenin signaling pathway inhibited by hGC33-null-NP or hGC33
hGC33-SFB-NP or hGC33 Attenuates HCC Cell Migration by Inhibiting Epithelial Mesenchymal Transition (EMT)
HCC is one of the deadliest cancers in the world, and its incidence is steadily increasing. Sorafenib is the only approved standard treatment for patients with advanced HCC, as it has been shown to improve the survival rate of these patients. However, clinical and preclinical observations suggest that sorafenib therapy has limited efficacy due to the rapid development of drug resistance. Therefore, elucidating the mechanism of escape resistance to sorafenib is a major emphasis in HCC research. In recent years, more and more attention has been paid to the role of epithelial mesenchymal transition (EMT) in the progress of HCC and the development of drug resistance. EMT refers to the transformation of epithelial to mesenchymal cells, which endows cells with the ability metastasize and invade, including acquisition of stem cell characteristics, reducing apoptosis and aging, promoting immunosuppression, and participating in the occurrence and development of cancer. The loss of E-cadherin expression is considered a key step in the carcinogenesis and EMT. EMT is a developmental multi-step molecular and cellular reprogramming process that cancer cells use to achieve aggression. This is mainly through down regulating the expression of E-cadherin, keratin, mucin, ZO-1 (tight junction protein); up regulating the expression of vimentin, alpha-smooth muscle actin (α-SMA), FN fibronectin, MMPs (degradation matrix), N-cadherin, snail, slug, twist, Rho, TGF-β, FGF, type I collagen, and type II collagen to achieve the invasion, metastasis, and anti-apoptosis of EMT characteristic tumor. The changes of these protein expressions mainly involve the activation of Wnt/β-catenin and Ras/Raf/MAPK signaling pathways.
Our experiments have shown that hGC33 antibody on the surface of NP vector can inhibit Wnt3a-induced β-catenin signal transduction, and then down regulate the expression of β—catenin, CD44, vascular endothelial growth factor (VEGF), cyclin D1, and c-MYC. Furthermore, hGC33-SFB-NP inhibits the activation of Ras/Raf/MAPK signal pathway and inhibits proliferation and apoptosis of HCC cells. hGC33 and SFB have a synergistic effect, inhibiting EMT and decreasing the migration of HCC cells (Fig. 8).

Effect of hGC33-SFB-NP on EMT inhibition. a Compared with the control group, the hGC33-SFB-NP treatment group had less cell migration. Photographs were taken under an optical microscope (magnification, × 200). The error value represents the standard deviation of three independent experiments. *Compared with the control group, p <0,01. b Compared with the control group, the EMT-related proteins snail, vimentin, and MMP-2 in HCCs treated with hGC33-SFB-NP decreased, whereas E-cadherin increased. c Molecular mechanism of EMT. EMT, epithelial–mesenchymal transition; MMP-2, matrix metalloproteinase-2; SFB, sorafenib; NP, nanoparticle
hGC33-SFB-NP Inhibits the Growth of Hepatocellular Carcinoma In Vivo and Improves the Survival Rate of Tumor-Bearing Mice
To evaluate the anti-tumor activity of hGC33-SFB-NP in vivo, HepG2 and Huh-7 cells were inoculated subcutaneously into the right abdomen and dorsal side of female BALB/c nude mice, respectively. When the tumor xenograft growth reached about 30 mm 3 , the mice were randomly divided into groups to further evaluate the inhibition of each group (hGC33-SFB-NP, hGC33-null-NP, SFB-NP, free hGC33, free SFB, and control group) HCC effect (n = 5 per group). It can be seen from Fig. 9a, b that hGC33-SFB-NP significantly slowed tumor growth in mice compared with the PBS control and other treatments. Compared with the PBS control, hGC33-null-NP, SFB-NP, free hGC33, and free SFB also had some inhibition of HCC, which is because free hGC33 and free SFB directly inhibit Wnt signal and Ras/Raf/MAPK, respectively. Such pathways can inhibit the proliferation of HCC cells to a certain extent. Although the nanoparticle-modified hGC33 (hGC33-null-NP) is connected to the nanosurface through chemical bonds, it did not affect hGC33′s targeting of GPC3 molecules and inhibition of Wnt activity. Nanoparticle-loaded SFB (SFB-NP), after being endocytosed by cells, was degraded to release SFB from the copolymer to inhibit the growth of HCC. In all, the inhibitory effect of hGC33-SFB-NP on HepG2 cell grafts was, as expected, more than on Huh-7 cell grafts, probably because HepG2 expresses GPC3 molecules.

The effect of hGC33-SFB-NP on xenotransplantation of HCC in nude mice and the changes of mice weight. Liver cancer cells were inoculated subcutaneously on the back of each nude mouse (n = 10). After 10 days, the tumor bearing mice were treated with PBS (control), free hGC33, free SFB, hGC33-null-NP, SFB-NP, and hGC33-SFB-NP. Tumor size (a , b ) and body weight (c , d ) of mice were monitored at designated time points
The body weight of nude mice in each treatment group also was measured, as shown in Fig. 9c, d. The body weight of the control group decreased gradually. The weight of mice in free hGC33, free SFB, SFB-NP, and hGC33-null-NP treatment groups also decreased progressively and not significantly less than in the control group. However, the weight of nude mice bearing HepG2 and Huh-7 treated with hGC33-SFB-NP only slightly decreased, and the weight remained relatively stable during the treatment cycle. These results support that the novel hGC33-SFB-NP nanodrug has no significant toxicity in nude mice, and the SFB loaded on the nanocarrier and the surface modified hGC33 can produce additive or even synergistic anti-tumor effect.
Discussion
To examine the suitability of hGC33-SFB-NP for targeted HCC therapy, we tested the model conjugates for their ability to bind to human glypican-3 on HCC cells in vitro; to inhibit glypican-3-positive HCC cell proliferation, migration, and Wnt/β-catenin signal transduction; and inhibit HCC that overexpress glypican-3 in vivo.
To covalently bind GPC3-specific antibody hGC33 with mal-PEG-b -PLGA nanoparticles, we cross-linked the free sulfhydryl group in the Fc segment of hGC33 with maleimide functionalized PEG-b -PLGA (mal-PEG-b -PLGA) by forming stable thioether bonds. Conjugation is a prerequisite for targeting of GPC3-positive HCC. A series of experiments, including the changes of nanoparticle diameter and zeta potential detected by lens and the intracellular uptake of hGC33-SFB-NP, verified the targeting of hGC33-SFB-NP to HepG2 (GPC3 + ) células. These results indicated that the binding activity of antibody hGC33 was not altered by the conjugation.
We directly detected the phagocytic effect of GPC3 + HepG2 and GPC3 − Li-7 cells on PEG-b -PLGA NP surface-modified hGC33 by confocal microscopy. After HepG2 and Li-7 cells were co-cultured with hGC33-coumarin 6-NP, the green signal intensity in HepG2 cells was significantly higher than in Li-7 cells, indicating that there were more nanoparticles in the HepG2 cells. This finding is consistent with the hGC33 antibody modified on the surface of PEG-b -PLGA NP specifically binding to glypican-3 on the surface of HCC cells and being internalized. The efficiency of hGC33-modified NP internalization depends on the expression level of GPC3 antigen on the cell surface.
We used the standard MTT assay to measure the efficiency of inhibiting the proliferation of hepatoma cells. Both hGC33-null-NP and hGC33 inhibited the growth of the GPC3-positive HCC cell line HepG2, but hGC33-null-NP and hGC33 did not affect the proliferation of GPC3-negative Li-7 cells (Fig. 3b). At the animal level, hGC33-null-NP or hGC33 alone inhibited the growth of Huh-7 and HepG2 xenografts to a certain extent, while hGC33-SFB-NP caused growth arrest of Huh-7 and HepG2 hepatoma xenografts in mice. The finding that hGC33-null-NP significantly inhibited GPC3-positive hepatoma cells showed that the inhibitory effect of PEG-b -PLGA NP surface-modified hGC33 on HCC cell proliferation depends on the expression of GPC3 antigen on the cell surface.
GPC3 regulates many pathways in HCC pathogenesis, including Wnt and YAP signaling [25,26,27]. GPC3 interacts with Wnt ligand and may be a coreceptor for Wnt and facilitate Wnt/Frizzled binding for HCC growth [28, 29]. We further examined the effect of nanodrug surface-modified hGC33 on Wnt signaling pathway in hepatoma cells. Like free hGC33, nanodrug surface-modified hGC33 inhibited the proliferation of hepatoma cells not only by blocking Wnt-induced signal transduction in HepG2 and Huh-7 cells of expressing GPC3, but also by inhibiting Wnt3a-induced β-catenin and YAP signal transduction. Previous studies have shown that YAP expression is regulated by β-catenin at the transcriptional level of HCC [30, 31]. In this study, free hGC33 and nanodrug surface-modified hGC33 inhibited Wnt3a-induced YAP activity, indicating that Yap/TAZ released from β-catenin complex can also initiate classic Wnt signaling transduction [32, 2]. These results indicate that typical Wnt and YAP cross talk through a variety of mechanisms. Compared with hGC33-null-NP and hGC33, hGC33-SFB-NP had stronger anti-proliferation and anti-migration ability in vitro and in vivo. Thus, hGC33 not only determines the specificity of HCC cells, but also increases the inhibitory effect of SFB on the proliferation and migration of HCC cells by blocking the key signals related to tumor growth, such as Wnt/β-catenin and Wnt/YAP signaling pathway.
Conclusion
Antibody hGC33 to glypican-3, a membrane protein that is overexpressed on hepatocellular carcinoma cells, increased binding of sorafenib-loaded polyethylene glycol-b-PLGA polymer nanoparticles (hGC33-SFB-NP) to glypican-3 on the cancer cells. Administration of the antibody-modified nanoparticles synergistically inhibited Wnt-induced signal transduction and Ras/Raf/MAPK signaling pathway; hepatocellular carcinoma cells were arrested in G0/1 phase by down-regulation of cyclin D1 expression, thus attenuating cancer cell migration by inhibiting epithelial–mesenchymal transition. hGC33-SFB-NP inhibited the growth of liver cancer in vivo and improved the survival rate of tumor-bearing mice.
Availability of data and material
Yes, all data have presented in the manuscript.
Abreviações
- AKT/PKB:
-
Protein kinase B
- c-MET:
-
HGFR:Hepatocyte growth factor receptor
- EE%:
-
Encapsulation efficiency %
- EMT:
-
Epithelial mesenchymal transition
- FTIR:
-
Espectroscopia de infravermelho com transformada de Fourier
- 1 H NMR:
-
1 H-nuclear magnetic resonance spectroscopy
- HCC:
-
Hepatocellular carcinoma
- LC%:
-
Drug loading %
- MAPK:
-
Mitogen-activated protein kinases
- PDI:
-
Índice de polidispersidade
- PI3K:
-
Phosphoinositide 3-kinase
- pRAD51:
-
Phospho-RAD51
- SFB:
-
Sorafenib
- TEM:
-
Microscopia eletrônica de transmissão
- YAP:
-
Yes-associated protein-1
MicroRNA exossômico derivado de macrófago M1-326 suprime a progressão celular do carcinoma hepatocelular por meio da via de sinalização NF-κB de mediação
Pesquisa sobre a preparação e características espectrais do grafeno / TMDs heteroestruturas
Nanomateriais
- Nanopartículas para terapia do câncer:progresso e desafios atuais
- Preparação e propriedades magnéticas de nanopartículas de espinélio FeMn2O4 dopadas com cobalto
- Nanopartículas como bomba de efluxo e inibidor de biofilme para rejuvenescer o efeito bactericida de antibióticos convencionais
- Efeitos sinérgicos de nanopartículas de Ag / BiV1-xMoxO4 com atividade fotocatalítica aprimorada
- Influência da rigidez elástica e adesão da superfície no salto de nanopartículas
- Avanços recentes em métodos sintéticos e aplicações de nanoestruturas de prata
- Toxicidade de nanopartículas de CoFe2O4 revestidas com PEG com efeito de tratamento da curcumina
- Efeito do agente de peptização ácida na relação anatase-rutilo e desempenho fotocatalítico de nanopartículas de TiO2
- Nanopartículas de ouro de formato redondo:efeito do tamanho da partícula e da concentração no crescimento da raiz de Arabidopsis thaliana
- Síntese Verde de Nanopartículas de Metal e Óxido de Metal e Seu Efeito na Alga Unicelular Chlamydomonas reinhardtii