


Efeito de nanopartículas projetadas na liberação de substâncias exopoliméricas do fitoplâncton marinho
Resumo
Nanopartículas projetadas (ENPs), produtos de nanotecnologias modernas, podem impactar potencialmente o ambiente marinho e representar sérias ameaças aos ecossistemas marinhos. No entanto, as respostas celulares do fitoplâncton marinho aos ENPs ainda não estão bem estabelecidas. Aqui, investigamos quatro espécies diferentes de diatomáceas ( Odontella mobiliensis , Skeletonema grethae , Phaeodactylum tricornutum , Thalassiosira pseudonana ) e uma alga verde ( Dunaliella tertiolecta ) para sua liberação de substâncias poliméricas extracelulares (EPS) sob o modelo de tratamentos ENP:dióxido de titânio de 25 nm (TiO 2 ), Dióxido de silício de 10-20 nm (SiO 2 ) e dióxido de cério de 15-30 nm (CeO 2 ) Encontramos SiO 2 ENPs podem estimular significativamente a liberação de EPS dessas algas (200-800%), enquanto TiO 2 A exposição a ENP induziu a liberação mais baixa. Além disso, o aumento do Ca intracelular 2+ a concentração pode ser acionada por ENPs, sugerindo que o processo de liberação de EPS é mediado por Ca 2+ vias de sinal. Com uma melhor compreensão do mecanismo celular mediado pela liberação de EPS induzida por ENP, potenciais medidas preventivas e de segurança podem ser desenvolvidas para mitigar o impacto negativo no ecossistema marinho.
Histórico
Nanopartículas projetadas (ENPs), que variam em tamanho entre 1 e 100 nm (em pelo menos uma dimensão), são usadas na fabricação de inúmeros bens de consumo, incluindo tintas e tintas para impressoras, detergentes, bactericidas, revestimentos, cosméticos, loções protetoras solares, pneus, construção de computador e entrega de medicamentos. Dada a aplicação promissora de ENPs, o financiamento para a National Nanotechnology Initiative (NNI) nos EUA sozinho se aproximou de US $ 1,4 bilhão em 2017 [1,2,3]. Estabelecer conhecimento básico em nanoescala foi o foco principal da comunidade de pesquisa em nanotecnologia na primeira fase. Em 2009, este novo conhecimento sustentou cerca de um quarto de um trilhão de dólares no mercado mundial, dos quais cerca de US $ 91 bilhões foram em produtos dos EUA que incorporam componentes em nanoescala [4]. Com o rápido desenvolvimento da nanotecnologia, é inevitável que os ENPs acabem chegando aos sistemas aquáticos. A principal preocupação com ENPs em termos de sua toxicidade potencial (por exemplo, o potencial para a produção de espécies reativas de oxigênio, ROS) no ambiente está relacionada à sua grande e única reatividade de superfície. No entanto, o impacto real no ecossistema marinho permanece amplamente desconhecido devido aos complexos fatores ambientais e biológicos das águas naturais e à variedade de PEVs [1, 5, 6]. Estudos anteriores demonstraram que os ENPs podem causar danos significativos ao ecossistema marinho baseado em algas [7, 8]. Os organismos marinhos (particularmente o fitoplâncton) têm demonstrado interagir com ENPs levando a repercussões negativas [9,10,11]. Com o potencial aumento da utilização da nanotecnologia em diversos campos, mais e mais ENPs podem entrar em ambientes aquáticos, de modo que as respostas celulares do fitoplâncton marinho aos ENPs exigem mais atenção [12,13,14,15,16,17,18,19,20, 21].
A maioria dos micróbios marinhos, sejam auto ou heterotróficos, são geralmente capazes de produzir substâncias exopoliméricas (EPS), que têm diversos papéis funcionais e propriedades físicas no ecossistema marinho atuando como inibidores de crescimento, promotores de crescimento, toxinas, eliminadores de metais ou como substratos para o ciclo heterotrófico [22,23,24,25,26]. EPS liberado de fitoplâncton e bactérias no oceano são biopolímeros coloidais aniônicos ricos em polissacarídeos que são críticos para a formação de géis marinhos, neve marinha e biofilmes, bem como para eliminação de colóides e oligoelementos e para fornecer proteção contra várias ameaças ambientais, incluindo ENPs [7, 15, 19, 20, 25, 27]. Além disso, acredita-se que a secreção de EPS seja uma resposta natural quando o fitoplâncton sofre vários tipos de estresse [8].
Ca 2+ é um segundo mensageiro comum envolvido em uma infinidade de vias de sinalização intracelular. Foi demonstrado que Ca 2+ é necessário para quimiotaxia, motilidade e adesão na diatomácea Amphora coffeaeformis [28]. Ca 2+ intracelular livre aprimorado níveis são conhecidos por levar à ativação da proteína quinase C, que está envolvida em muitas vias de sinalização intracelular [29]. Uma vez que o lançamento de EPS está intimamente relacionado à motilidade e adesão das diatomáceas, foi proposto que um Ca 2+ - o processo de secreção mediado controla a liberação de EPS das diatomáceas [30], e a evidência direta verificando Ca 2+ sinalização, exocitose e correlação de Ca 2+ sinalização com exocitose foi relatada em nosso estudo anterior [31]. Estudos anteriores também demonstraram que as interações com ENPs podem alterar o Ca 2+ intracelular vias, que são essenciais para a sinalização celular [29, 32,33,34]. Ca intracelular específico 2+ as mudanças na concentração são importantes nos processos de sinalização e secreção celular; no entanto, não há relatos de dióxido de titânio (TiO 2 ), dióxido de silício (SiO 2 ), ou dióxido de cério (CeO 2 ) para alterar Ca intracelular 2+ nível no fitoplâncton.
Em 2013, Quigg et al. [8] resumiu os efeitos tóxicos diretos e indiretos dos ENPs nas algas. Em nossos experimentos anteriores, os ENPs mostraram facilitar a agregação de EPS [35]. Nesse sentido, os EPS podem exacerbar ou reduzir a toxicidade direta induzida por ENP para organismos aquáticos [7, 15, 36]. No entanto, a medição direta da liberação de EPS do fitoplâncton sob estresse ENPs nunca foi relatada. Neste estudo, o objetivo é estudar a liberação de EPS de quatro espécies diferentes de diatomáceas ( Odontella mobiliensis , Skeletonema grethae , Phaeodactylum tricornutum , Thalassiosira pseudonana ) e uma alga verde ( Dunaliella tertiolecta ) em tratamentos ENP. Ao compreender os mecanismos subjacentes da liberação de EPS induzida por ENP no fitoplâncton, a implementação de medidas preventivas e de segurança pode mitigar os efeitos potencialmente prejudiciais para os organismos marinhos.
Resultados e discussões
Caracterização ENP
Espalhamento de laser dinâmico (DLS) foi usado para caracterizar métricas de tamanho dos seguintes ENPs suspensos em água pura:TiO 2 , SiO 2 e CeO 2 . A distribuição do tamanho de partícula variou de 7 a 66 nm em TiO 2 , 9 a 66 nm em SiO 2 , e 12 a 70 nm em CeO 2 . Alguns tamanhos maiores podem ser devido à agregação ou aglomeração, enquanto o tamanho predominante para TiO 2 é 25 nm, SiO 2 é de 10 a 20 nm, e CeO 2 é de 15 a 30 nm, que são consistentes com as informações do fabricante (Fig. 1).
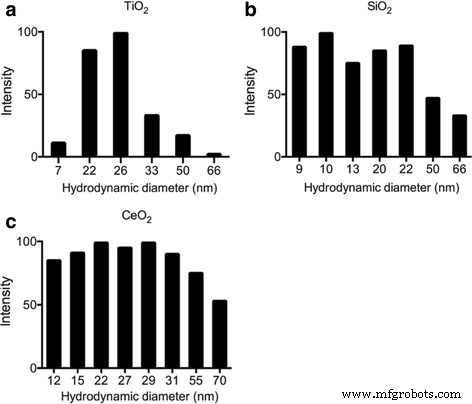
Caracterização ENP por avaliação DLS de a TiO 2 , b SiO 2 , e c CeO 2 no meio L1 após sonicação mostrando sua distribuição de tamanho. A concentração final de ENP na amostra DLS é de 1 μg / ml, o tempo de medição é de 3 minutos logo após a sonicação
ENPs induzem Ca intracelular 2+ Concentração no fitoplâncton
Para investigar se os ENPs podem induzir um aumento no Ca intracelular 2+ concentração, as células do fitoplâncton (OD 600 =0,8) foram carregadas com corante Fluo-4AM e expostas a 1 mg / ml de TiO de 25 nm 2 , 10–20 nm SiO 2 e 15-30 nm CeO 2 ENPs respectivamente. A mudança no Ca intracelular 2+ a concentração, representada pela intensidade de fluorescência nas células do fitoplâncton, foi monitorada por 150 s. A Figura 2a-e mostra que 1 mg / ml de três respectivos ENPs aumentaram Ca 2+ concentração em SiO 2 em aproximadamente 50–300%, TiO 2 em aproximadamente 40%, e CeO 2 em aproximadamente 150–200%, enquanto as condições de controle (meio L1) permaneceram inalteradas. Os resultados mostram que ENPs podem induzir Ca intracelular significativo 2+ respostas no fitoplâncton e sugerem que o fitoplâncton responde a ENPs distintos por meio de Ca 2+ vias de sinalização. Nossos dados indicam apenas pequenas alterações no Ca 2+ intracelular níveis quando TiO 2 está presente, potencialmente atribuído à morte substancial de células do fitoplâncton por TiO 2 - toxicidade induzida [37, 38]. Em nosso estudo anterior, TiO 2 aumento solicitado no Ca intracelular 2+ concentração [34] ao lado de apoptose celular significativa [39]. No entanto, SiO 2 surpreendentemente mostrou o Ca 2+ intracelular mais óbvio aumento para todas as espécies de fitoplâncton, enquanto CeO 2 só pode desencadear um Ca intracelular intermediário 2+ aumento da concentração. Pesquisas anteriores sugeriram potencial de alto CeO 2 concentrações (> 50 mg / ml) para induzir estresse oxidativo intracelular e elevação do Ca intracelular 2+ níveis, embora os efeitos fossem pequenos, e apoiaram nossa descoberta [40]. Também medimos o potencial zeta de cada ENPs na água do mar artificial para abordar o efeito potencial que a carga superficial pode causar; no entanto, o valor era baixo. A medição indicada os ENPs são considerados aproximadamente neutros [41] (arquivo adicional 1:dados suplementares). Isso serviu como o primeiro relatório em que ENPs díspares foram encontrados para induzir Ca intracelular 2+ mudanças de concentração em fitoplâncton específico, em última análise, pavimentando um novo caminho para pesquisas futuras.
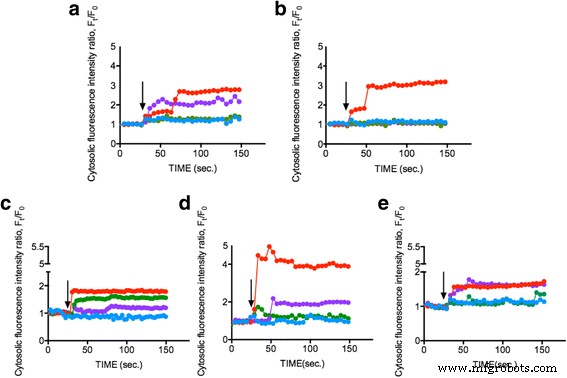
Medição de Ca intracelular 2+ concentração após estimulação por diferentes ENPs. Diferentes células do fitoplâncton a Dunaliella tertiolecta , b Thalassiosira pseudonana , c Skeletonema grathae , d Phaeodactylum tricornutum , e e Odontella mobiliensis foram tratados com TiO 2 25 nm (verde), SiO 2 10–20 nm (vermelho), CeO 2 15-30 nm (roxo) com uma concentração de 1 mg / ml e controle (azul). A seta preta indica o momento em que os EPNs foram aplicados (30 s). As medições mostram dados representativos de uma média de 20 células individuais
Liberação de EPS induzida por ENP no fitoplâncton
O ensaio de lectina ligada à enzima (ELLA) foi usado para avaliar a quantidade de liberação de EPS a partir de células fitoplanctônicas quando estimuladas com TiO 2 , SiO 2 e CeO 2 ENPs, faixa de concentração de 1 μg / ml a 5 mg / ml com base em estudos anteriores para TiO 2 [42, 43] e CeO 2 [44,45,46]. A secreção de EPS foi normalizada para a quantidade total de DNA do fitoplâncton (arquivo adicional 1:dados de suplemento) para ter uma base igual para comparação. Comparado com o controle, descobrimos que 10-20 nm SiO 2 é capaz de aumentar a liberação de EPS em até 550% em Dunaliella , 500% em Thalassiosira , 1000% em Esqueletonema , 400% em Odontella , e 900% em Phaeodactylum (Fig. 3). Quando as espécies de fitoplâncton foram expostas ao TiO 2 , não houve efeito forte na secreção de EPS, pois apenas Skeletonema e Phaeodactylum mostraram mudanças significativas. Os dados de liberação de EPS são, portanto, consistentes com nosso Ca 2+ intracelular resultados de concentração. TiO 2 não apresentou impacto significativo na produção de EPS, semelhante ao fato de Ca +2 intracelular as concentrações mostraram mudanças muito limites devido à toxicidade do TiO 2 para o fitoplâncton. A produção e resíduos de ROS podem levar a muitas complicações como apoptose no fitoplâncton [47,48,49]. No CeO 2 tratamento, os resultados mostraram um efeito menor em Dunaliella , Esqueletonema , Odontella e Phaeodactylum . No entanto, SiO 2 mostrou a indução de EPS mais significativa em Thalassiosira pseudonana (cerca de 600%) e Skeletonema grethae (cerca de 1000-1500%). Esses dados indicam que diferentes ENPs podem induzir a liberação específica de EPS do fitoplâncton e Ca intracelular 2+ as alterações também correspondem aos resultados da liberação do EPS. Avaliando as mudanças no Ca intracelular 2+ concentração, é evidente que há uma conexão direta no Ca 2+ vias celulares nas quais os ENPs evocam a secreção de EPS do fitoplâncton. A observação aqui está de acordo com nossos estudos anteriores baseados em Phaeocystis Lançamento do EPS [31]. Os resultados fornecem evidências diretas de que o fitoplâncton pode detectar e distinguir ENPs respondendo com diferentes liberações de EPS reguladas por Ca 2+ vias celulares.
O uso de ELLA nos permitiu determinar a liberação de EPS por meio das interações do fitoplâncton com os ENPs. Nossos resultados indicam que a secreção de EPS foi significativamente aumentada conforme o fitoplâncton interagia com SiO 2 para Dunaliella tertiolecta , Thalassiosira pseudonana e Skeletonema grethae . Parece que essas diatomáceas são preparadas para reconhecer SiO 2 partículas. No entanto, em Phaeodactylum tricornutum , uma forte secreção de EPS não foi encontrada. Esta diferença representa a liberação de EPS desencadeada por ENPs dependentes das espécies de fitoplâncton e concentração de ENPs (Fig. 3). Em um estudo anterior, derramamentos de óleo causaram grandes liberações de EPS microbiano marinho que foram propostas para neutralizar as consequências negativas dos derramamentos de óleo [50]. Além disso, Boglaienko e Tansel descobriram que SiO 2 partículas foram capazes de remover agregados de óleo de forma eficiente [51]. Nossa descoberta fornece um novo mecanismo potencial em que SiO 2 de baixa toxicidade partículas podem induzir a liberação de EPS de fitoplâncton específico, potencialmente facilitando a remoção do derramamento de óleo ao promover a agregação de EPS. Nunca foi relatado que o dióxido de cério perturba os ecossistemas marinhos baseados no fitoplâncton. Os resultados aqui mostraram CeO 2 Os ENPs podem afetar todo o fitoplâncton aqui, exceto Thalassiosira pseudonana. CeO 2 ENPs podem, como SiO 2, têm a capacidade de aumentar a liberação de EPS de um fitoplâncton específico para aplicações de mitigação de óleo.
Conclusões
A interação PEV-ambiente marinho está se tornando cada vez mais crítica devido às descargas atuais e futuras de nanomateriais. Aqui, demonstramos a secreção aumentada de EPS como um dos principais efeitos dos ENPs para o fitoplâncton. Também fornecemos evidências de que fitoplâncton diferente pode responder de forma diferente a vários estresses ENP, regulando Ca 2+ caminhos. No entanto, uma avaliação completa dos PEV para o ecossistema marinho necessitaria de mais investigações para fornecer conhecimento detalhado e compreensão das interações entre os nanomateriais e os organismos marinhos.
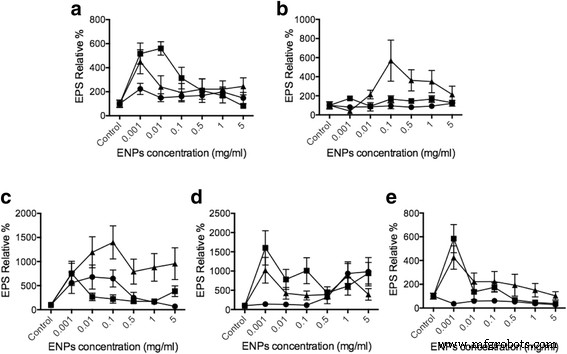
Liberação de EPS acionada por vários ENPs. Diferentes células do fitoplâncton a Dunaliella tertiolecta , b Thalassiosira pseudonana , c Skeletonema grathae , d Phaeodactylum tricornutum , e e Odontella mobiliensis foram tratados com TiO 2 (círculos), SiO 2 (triângulos), CeO 2 (quadrados), respectivamente, com concentrações de 5 mg / ml e 1 mg / ml, 0,5 mg / ml, 0,1 mg / ml, 10 μg / ml, 1 μg / ml ( n =3)
Métodos
Cultura do fitoplâncton
Culturas em lote de Odontella mobiliensis (CCMP597), Dunaliella tertiolecta (UTEX999), Skeletonema grethae (CCMP775), Phaeodactylum tricornutum (UTEX646), Thalassiosira pseudonana (Provasoli - coleção de cultura de fitoplâncton marinho de Guillard, West Boothbay Harbor, MN, EUA) foram cultivadas em meio marinho L1 (Sigma, MO, EUA) em um ciclo de 14:10 (claro:escuro) a 100 μmol m −2 s −1 e 24 ° C em condições axênicas. A fase de crescimento da cultura foi determinada por contagem de células em hemocitômetro.
Nanopartículas e caracterização
Todos os ENPs, TiO 2 , SiO 2 , CeO 2 (Sigma-Aldrich, MO, EUA), foram sonicados em água pura antes do uso. Os ENPs foram reconstituídos com meio L1 filtrado (Sigma, MO, EUA) antes de serem testados. O tamanho dos ENPs foi confirmado independentemente usando espalhamento de laser de dinâmica homodínica (DLS). Resumidamente, as amostras de água do mar foram filtradas novamente através de uma membrana Millipore de 0,22 μm (pré-lavadas com HCl 0,1 N) e vertidas diretamente em cinco células de dispersão de 10 ml que foram posicionadas no goniômetro de um espectrômetro de laser Brookhaven BI-200SM (Brookhaven Instruments, NY, EUA). A função de autocorrelação das flutuações de intensidade de espalhamento detectadas em um ângulo de 45 ° foi processada em linha por um autocorrelador Brookhaven BI 9000AT, e a distribuição do tamanho de partícula foi calculada pelo método CONTIN (Provencher, 1982). Os resultados de cada amostra foram coletados em triplicata logo após a sonicação. A calibração do espectrômetro DLS foi realizada usando suspensões padrão de microesferas de látex monodispersas (Polysciences, PA, EUA).
Tratamento ENP
As células do fitoplâncton foram cultivadas em uma placa de 96 poços com meio L1 por 24 h. As células foram tratadas com estoques de ENP:5 mg / ml e 1 mg / ml, 0,5 mg / ml, 0,1 mg / ml, 10 μg / ml, 1 μg / ml do TiO 2 , SiO 2 e CeO 2 (Sigma-Aldrich, MO, EUA) ou meio L1 (controle) por 48 h. O sobrenadante contendo EPS segregado foi coletado e brevemente centrifugado a 4000 rpm para remover os ENPs residuais. Este protocolo foi adaptado de nossa publicação anterior [34]. O intervalo de concentração usado aqui não se destina a representar ou imitar os níveis atuais de ENP no meio ambiente, mas visa avaliar o impacto potencial total dos ENPs no fitoplâncton marinho e investigar os mecanismos celulares associados. Como um nanomaterial emergente promissor, os ENPs ainda não atingiram sua capacidade comercial plena. A avaliação detalhada de seus impactos ecológicos completos é muito necessária antes que os ENPs entrem no mercado de produtos domésticos e comerciais para introduzir mais ENPs no oceano.
Ensaio de lectina ligada a enzima (ELLA)
O sobrenadante contendo o polissacarídeo secretado foi coletado e brevemente centrifugado a 1700 rcf (Megafuge 1.0R) para remover os ENPs residuais. O sobrenadante foi então incubado em uma placa de 96 poços (Nunc MaxiSorp, VWR, CA, EUA) durante a noite a 4 ° C. Posteriormente, a placa de 96 poços foi lavada com PBST (PBS + Tween-20 a 0,05%) e PBS e depois bloqueada com BSA a 1%. A placa de 96 poços foi lavada novamente com PBST e PBS e incubada com lectina (Concanavalin A, ConA) (Sigma-Aldrich, MO, EUA), conjugada com peroxidase de rábano (HRP; 5 mg / ml) (Sigma-Aldrich, MO , EUA), a 37 ° C durante 1 h. O substrato, 3,39,5,59-tetrametilbenzidina (TMB; Sigma-Aldrich, MO, EUA), foi adicionado a cada poço em temperatura ambiente seguido por H 2 SO4 (Sigma-Aldrich, MO, EUA) para encerrar a reação. A densidade óptica foi medida a 450 nm por PerkinElmer VICTOR3 (MA, EUA). Este protocolo foi adaptado de nossa publicação anterior [34, 52].
Determinação de DNA
O pellet contendo fitoplâncton foi coletado e obtido o kit ZR-96 Quick-gDNA (ZYMO Research, CA, EUA). Em resumo, o tampão de lise 4 × foi usado para quebrar as células do fitoplâncton e fluir através da coluna de ligação de DNA, eluída por tampão de eluição no final. As concentrações de DNA foram medidas por NanoDrop ND-1000 (Thermo, CA, EUA). O protocolo foi adaptado do protocolo do kit fabricado.
Medições de Ca intracelular 2+ Concentrações induzidas por ENPs
As células do fitoplâncton foram então carregadas com um corante Fluo-4AM (1 mM) (Kd =335 nM, λEx =494 nm e λEm =506 nm, ThermoFisher, CA, EUA) por 60 min [31]. Após o carregamento do corante, as células do fitoplâncton foram enxaguadas, incubadas com meio L1 e tratadas com 1 mg / ml de TiO 2 , SiO 2 e CeO 2 respectivamente. Todos os experimentos de sinalização de cálcio foram realizados em um microscópio Nikon (Nikon Eclipse TE2000-U, Tóquio, Japão). O protocolo e as condições foram adaptados de publicações anteriores [31, 34].
Potencial Zeta de medição ENP
Para medir as cargas superficiais dos ENPs, o potencial zeta (ζ) dos ENPs foi medido com um Zetasizer Nano ZS, Malvern, na presença de água do mar artificial a 25 ° C. Depois que os dados foram coletados de cada amostra, os valores registrados foram calculados.
Análise estatística
Os dados são relatados como médias ± DP. Cada experiência foi realizada de forma independente pelo menos três vezes. Os histogramas foram feitos por GraphPad Prism 6.0. (GraphPad Software, Inc., San Diego, CA, EUA).
Uma metodologia de duas etapas para estudar a influência da agregação / aglomeração de nanopartículas no módulo de Young de nanocompósitos poliméricos
RGO-PEDOT híbrido tratado com ozônio por UV:PSS como um material de transporte de furo eficiente em células solares de perovskita planar invertida
Nanomateriais
- Nanopartículas de semicondutor
- Nanopartículas plasmônicas
- O efeito do pH em corantes amarelos do jardim
- Mecanismo de formação de superredes bem ordenadas de nanopartículas densamente compactadas depositadas da fase gasosa em superfícies livres de modelo
- Nanopartículas como bomba de efluxo e inibidor de biofilme para rejuvenescer o efeito bactericida de antibióticos convencionais
- Bioflavonóide direcionado ao receptor de folato Nanopartículas de quitosana carregadas com genisteína para efeito anticâncer avançado em cânceres cervicais
- Nanotecnologia:do Sistema de Imagem In Vivo à Entrega Controlada de Medicamentos
- Efeito do método de síntese de La1 - xSr x MnO3 nanopartículas de manganita em suas propriedades
- Toxicidade de nanopartículas de CoFe2O4 revestidas com PEG com efeito de tratamento da curcumina
- Efeito do agente de peptização ácida na relação anatase-rutilo e desempenho fotocatalítico de nanopartículas de TiO2